Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#601 05 December 2022 14:09:41
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
По мелким котам – Felis…
В плиоцене Северной (Марокко) и Восточной (Кения, Танзания) Африки с датировкой 4-2.5 млн фиксируются интересные образцы мелкоразмерной кошки:
https://www.researchgate.net/publicatio … Casablanca
https://www.researchgate.net/publicatio … hern_Kenya
https://www.researchgate.net/publicatio … _Carnivora
Братцы, это таксон родового ранга. Вовсе не рысь, но ещё и не Felis как таковой.
Мандибула архаичная – ближе к сервалу и робустная, на р3 парастилид и метастилид почти не выражены, метастилид на р4 возвышен (естессно, редуцирован). Апоморфными признаками более прогрессивных филогенетических линий здесь почти «не пахнет».
Это на бифуркационной точке, когда протофелисы отделились от группы Prionailurus, но радиировать в Азию и Европу ещё не начали, примерно 3.5-3 «ляма». Аппендиксные формы из южноафриканского местонахождения Cooper’s D и израильской Ubeidiya датируются ранним плейстоценом - 1.5 млн, но образуют единую морфогруппу с плиоценовым материалом из северо-восточной Африки. Двестипудово это один род, причём новый.
Kryptofelis
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#602 01 July 2023 19:27:51
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Очень забавны американские мелкие кошки: оцелот, маргай, онцилла и пр. Композиционно они несколько однородны и, действительно, эндемичны. Кроме того, несколько самобытны морфологически, потому что краниодентально выглядят как нечто среднее между рысеподобными котами и архетипичными фелинами. Также имеются посткраниальные адаптации в строении костно-мышечного аппарата (задних конечностей) и, соответственно, локомоции.
Заметна некоторая схожесть с группой Pardofelini (Pardofelis, Catopuma ), но это, скорее, конвергентность.
Согласно последним данным (Kitchener et al., 2017) на генетической основе всех объединяют в род Leopardus . НО!....
Однозначно, в этой группе существуют морфологические различия, и, в приватном воззрении, эти исследования указывают лишь на генетическую дифферентность (филогеографическую структуру) среди современных популяций, в связи с чем, возможно, Leopardini лишь включает комплекс недавно дивергировавшихся форм, чья нынешняя генетическая взаимосвязанность и экологическая дифференциация до сих пор окончательно не урегулирована (Oliveira 1998; Sousa, Bager, 2008; Trigo et al,. 2008).
Парадоксально, что самые ранние фоссильные находки датируются, на удивление, очень поздно: vorohuensis Berta, 1983 – средний плейстоцен Ю. Америки: энсенадий Аргентины; 0.7-0.5 млн. близкой к предковой линии оцелотов (Seymour, 1999; Prevosti, 2006; Johnson et al., 2006).
В Северной Америке - amnicola Gillette 1976 – поздний плейстоцен. Голотип и паратипы Leopardus amnicola включают, в основном, фоссильный дентомандибулярный материал. Датировка – поздний плейстоцен, ранчолабрий (Ray et al., 1963; Kurten, 1965; Werdelin, 1985; Morgan, Emslie 2010; Smith et al., 2016) и данная форма интерпретируется как отдельный таксон, близкий к апоморфным образцам Leopardus.
Генетические исследования всё же не подтверждают окончательный вердикт одного вида. Поэтому группу целесообразно ранжировать на подроды Leopardus, Oreailurus и Oncifelis (включая Lynchailurus).
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#603 26 November 2023 22:59:43
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Коллеги, факультативные изыскания по Miracinonyx.
В настоящее время приняты два вида: ранний (inexpectatus) и поздний (trumani). Но если с позднеплейстоценовым/ранчолабрейским trumani более-менее определённо (есть некоторые морфометрические отличия в сангамонии и висконсине, но там выборки небольшие), то с inexpectatus несколько вопросительно. Дело в том, что его таксономическая история включает ряд синонимических значений (Felis longicrus, Smilodontopsis mooreheadi и пр.), одно из которых действительно заслуживает внимания: studeri .
Временной диапазон Miracinonyx inexpectatus охватывает ранний и средний плейстоцен Северной Америки: от 2.5. млн до 300 тлн. На самом деле странно, что за два «ляма» столь морфопластичная форма как кошка и специализированная в тренде «гепардоподобия» не менялась.
Голотип studeri (Savage 1960) датируется бланком, inexpectatus (Cope, 1895) – ирвингтонием. Дедушка Куртен (Kurtén, 1976. Fossil puma (Mammalia: Felidae) in North America) объединил ранне- и среднеплейстоценовых мирацинониксов (тогда он считал их аберрантными формами пум) на основе исключительно морфометрики. И на протяжении почти полувека так и считается…
Работ по морфологической дифференции бланкских и ирвингтонских мирацинониксов не знаю. Но отличия наглядны. По мандибулам, вверху – studeri (WT 1218, Техас), ниже, в сравнении, – образчик inexpectatus из Флориды: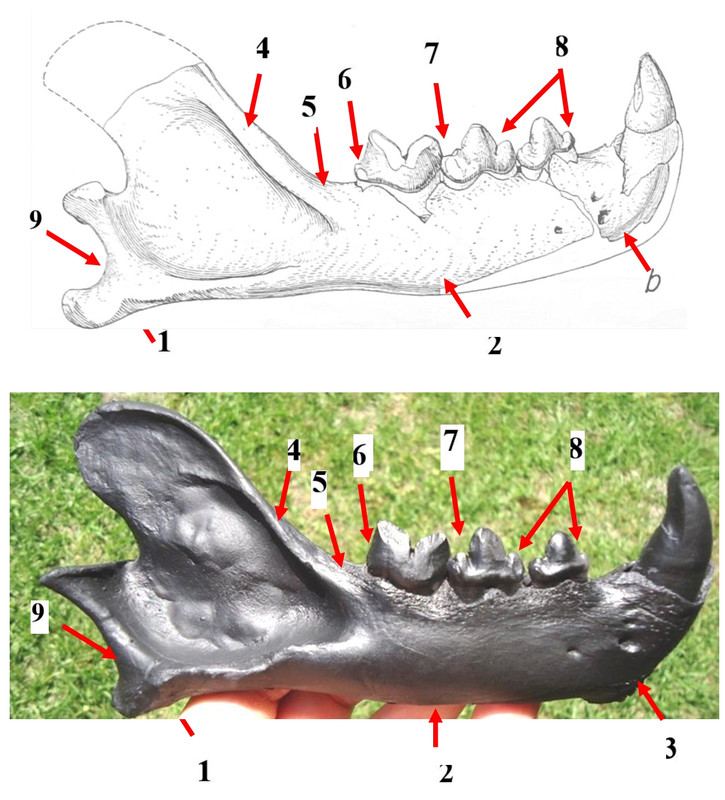
1 - угловой отросток нижней челюсти удлинён и является продолжением карнассиальной линии (менее загнут)
2 - тело нижней челюсти имеет изгиб в области р3/4 в отличие от спрямлённой вентральной линии
3 - подбородочная область (симфизеальная область) заметно менее вертикализированная
4 - гребень венечного отростка более утолщённый
5 - вертикальная ветвь, восходящая от клювовидного отростка, демонстрирует не столь крутой подъём от m1
6 - талонид-метаконидный комплекс на m1 выражен
7 - метастилид на р4 снижен, задний дополнительный бугорок хорошо обозначен
8 - парастилиды на р3/4 более крупные
9 - мыщелковая вырезка более углублённая
Есть также отличия по черепу и верхним зубам: мозговая коробка у studeri более удлинённая, височное сужение менее обозначено, височная линия постериорной области лобной кости более заострённая, затылочный гребень менее выражен и затылочная область более расширена, направленность верхнего клыка более антериорная, протокон на Р4 заметно укрупнён, паракон более заострён, выемка между параконом и метастилем явно выражена.
Бланский (2.5-1.8 млн) Miracinonyx studeri размерно и ещё во многих морфопризнаках сходен с ранними пумаподобными котами, лишь в ирвингтонии приобретя номинальный габитус. Но в приватном воззрении он всё же достоин, как минимум, подвидового статуса.
P.S. Ранне- и позднеирвингтонских мирацинониксов тоже можно было бы ранжировать. Но там разница в промерах обезображена, очевидно, половым диморфизмом; и, вообще-то, лучше «вживую» с образцами общаться.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#604 22 July 2024 20:08:05
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Наконец-то сам для себя распутал эволюционно-филогенетический узел пумагепардовых кошек Северной Америки.
Вообще, эта триба, таксономически, наверное, самая продуктивная в Felinae, породившая размерно самых терминальных представителей подсемейства: пум и гепардов. Причём, на удивление, на всех трёх материках независимо. Пумаморфы (из Laetoli, Woranso-Mille, Omo Shungura C-Usno) и, собственно гепарды (Acinonyx) - в Африке; в Евразии - пумаподобный Viretailurus и гепардоподобные Abacinonyx, Sivapanthera; в Сев.Америке - Miracinonyx и номинальная Puma.
Именно пумагепарды Неарктики представляли самое больное место… Ещё с времён 2-й мировой (Pocock, 1940; Simpson, 1941; Goldman, 1946), затем в лохматых 70 гг. (Morgan, Seymour 1976; Kurtén, 1976) и в XXI веке (Culver et al., 2000; Wozencraft, 2005; Hodnett et al., 2022) признавалось , что это не эндемики, а мигранты из Старого Света. Вот только имелись нестыковки.
Разумеется, пума и мирациноникс - близкородственные виды. Последнего до начала 80-х именовали то Puma, то Acinonyx, а признание Miracinonyx как род лишь в 90-х было общепризнано. В то же время мирациноникс распространён в Северной Америке от раннего до позднего плейстоцена, тогда как пума известна лишь со среднего.
Действительно, род Puma фиксируется в Новом Свете только 0.7 млн: начало позднего ирвингтония в Северной Америке и энсенадий в Южной. Всё это прекрасно согласуется с образцами пумаподобных котов из северо-восточной Азии, которые в среднем плейстоцене наряду с ягуароподобными пантерами преодолели Берингию и начали победный конкистадорский марш по Америкам.
Кстати, ранние пумы (североамериканская bituminosa и южноамериканская platensis) не сильно отличаются от современной concolor, которая сформировалась после висконсинского оледенения (30 тлн). В основном, примечательны более крупными (15-20 %) размерами и в этом пума повторяет сценарий ягуара по освоению Нового Света, вплоть до реверсивности матримональных популяций.
Но если мирациноникс фиксируется с раннего плейстоцена, а пума – на целый «лям» позже, то откуда он вообще взялся? Потому что самый примитивный мирационикс (studeri) – бланко, датируется примерно 2 млн, причём – Техас!!! Т. е. эволюция уже состоялась и ареал таксона занимал почти весь материк. В то же время крупных пумаподобных кошек в северо-восточной Азии (откуда могла быть «подпитка») ещё нет, а гепардоподобная Sivapanthera на роль предковой формы вряд ли подходит вследствие более высокой краниодентальной специализации, чем у ранних мирацинониксов.
И тут самое время вспомнить о позднеплиоценовой Puma lacustris. Примитивная пумаподобная кошка размером с крупную рысь или небольшую самку пумы. Ввиду сравнительной морфологии (несколько архаичной), датировки (поздний плиоцен – ранний плейстоцен) и местонахождений (центральная часть Северной Америки) как предковая форма и Miracinonyx и Herpailurus (ягуарунди) даже очень и очень...
Но здесь возникают две проблемы:
1. Предположительно, lacustris мигрирует в Северную Америку из Азии в конце плиоцена наряду с ранней рысью (Lynx rufus), гомотерием (Homotherium ischyrus) и оцелото-маргаями (Leopardini). Исход истинной Puma гораздо позже: вместе с ягуароподобными котами, а также поздними формами рыси, гомотерия и пещерной пантерой (atrox). Только в Азии подобных находок из плиоцена нет.
2. При более подробном и аргументированном рассмотрении данной гипотезы Puma lacustris следует считать уже не пумой, а таксоном нового рода (как и пумаподобных кошек из плиоцена Африки).
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#605 29 July 2024 14:59:26
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Вот не даёт покоя «озёрная» пума…
† Puma lacustris Gazin 1933 – озёрная кошка
Felis lacustris Gazin 1933, р. 251; Lynx lacustris Savage 1960, р. 324; Felis lacustris Bjork 1970, р. 39; Kurten, Anderson 1980, р. 195; Werdelin 1985, р. 201; Lynx lacustris Hulbert 1992, р. 19; Felis lacustris Seymour 1999, р. 433; Puma lacustris Morgan, White 2005, р. 123; Ruez 2009, р. 57.
Этимология. (лат.): lacustris – «озёрная» (именование основывается на старом геологическом названии оз. Хагерман – остатки были обнаружены среди отложений древнего озера).
Голотип. USNM V 12611 – фрагмент правой горизонтальной ветви нижней челюсти с клыком и зубным рядом р3-m1, национальный музей естествознания (Smithsonian Institution, National Museum of Natural History), Вашингтон, США.
Местонахождение голотипа. Хагерман (Hagerman), Айдахо; плиоценовый озёрный горизонт, 3.2 млн.
Другие местонахождения. Айдахо (Little Jacks Creek, Sugar Creek, Grand View); Аризона (Curtis Ranch); Вашингтон (Taunton); Калифорния (Vallecito Creek); Флорида (Upper Bone Valley, 111 Ranch); Техас (Overton, Cita Canyon, Red Quarry, Marmot Quarry); Мексика (Miraflores); бланко, 3.5-2.6 млн.

Фото голотипа, USNM V 12611.
Дифференцированный диагноз и аналитические выводы приводить не буду. Информация имеется в свободном доступе. Ищущий да обрящет… Вкратце изложу, что размеры крупнее рыси, но меньше пумы. Зубной ряд массивный, клыки короткие и укрупнённые, премоляры антериорно сужены, карнассиальные зубы небольшие. Нижняя челюсть с неглубоким симфизом, имеющим закруглённый передний край, не выступающий вниз. Конечности относительно короткие и робустные. Передние пропорциональны рысеподобным кошкам, задние имеют тенденцию к увеличению и в пропорциях ближе к пуме, но грацильнее.
Ниже собственная реконструкция черепа на основе наиболее полных образчиков и внешнего вида. Уж не взыщите, за карандаш давно не брался. 
Нарекаю ![]() сию кошку LESTAILURUS (др.-гр.): lest-(es) – «хищник»: (лат.): -ailurus «кошка». Потом посмотрим…
сию кошку LESTAILURUS (др.-гр.): lest-(es) – «хищник»: (лат.): -ailurus «кошка». Потом посмотрим…
Краткое приватное мнение – явно отличается от ранних рысеподобных североамериканских кошек (Lynx rufus=rexroadensis) как краниодентально, так и посткраниально. В сравнении с Miracinonyx и Puma также заметны морфологические отличия, как минимум, в зубных особенностях (более узкие премоляры с пропорционально массивными карнассиалами), а также отличительная ориентация сочленовных фасеток костей конечностей и их общая форма в различных особенностях, которые демонстрируют срединные характеристики промеров между рысями и пумами (передне-задние диаметры, поперечные и пр.).
Вывод. Это вовсе не архаичная форма пумы, а отдельный род пумаподобных фелин. Во всяком случае, весьма претендует на роль предка Miracinonyx.
P. S. В личном воззрении ягуарунди, Herpailurus, также - всё же отдельный род, а вовсе не Puma. С таким отъявленно-фанатичным подходом можно и мирациноникса к пуме причислить.
P. P. S. Очень странно, что при описании ископаемой ягуарунди южноамериканские псевдоколлеги (Chimento et al., 2014. Puma (Herpailurus) pumoides (Castellanos, 1958) nov. comb. ) её вообще не упоминают. Даже вскользь…
А теперь, братцы, риторические вопросы, которые не лишены эмпирики.
Извините за «я»-канье, но я не сторонник как чрезмерного объединения, так и оголтелого дробления ископаемых таксонов. К примеру, для меня пещерный лев это - пещерная пантера (Panthera spelaea) и к современному льву (P. leo) отношение имеет лишь сестринское. Разумеется, это очень близкородственные формы, но виды ведь разные. Также как у тигров: тринильская пантера P. trinilensis это ещё не совсем тигр (P. tigris), а в леопардах Panthera incurva - это ещё не леопард (P. pardus).
Аналогично можно экстраполировать и на трибу пумагепардовых кошек. Гипотетически в среднем-позднем плиоцене Азии появилась группа крупноразмерных котов, инициированная общим похолоданием (финал пьяченция (прикольная фонетическая этимология) и граница плиоцен-плейстоцена), что знаменуется революционной перестройкой фаунокомплексов. В гильдии фелид махайродовые начинают постепенно сдавать позиции, а фелины наращивают мускулы.
Затем грянул климатический апокалипсис, который продолжался несколько тысячелетий, в результате которого саблезубые кошки дифферентно адаптировались на лесные и лесостепные морфотипы (в Голарктике Homotherium и Megantereon), а место универсальных доминантов заняли представители подсемейства Felinae (хронологически ненадолго, всего пол-ляма до воцарения пантер).
Вот здесь победным маршем по всей планете идут пумаподобные коты (от которых в Азии, Африке и Сев. Америке независимо эволюционировали гепардоподобные).
Г-н Хэммер их всех объединяет под флагом Puma. Что представляется не совсем верным. Архетипом выглядит наиболее изученный Viretailurus, которого дядюшка Гельмут обозвал европейской пумой (Puma pardoides). Но ведь это – лишь европейский вектор распространения пумаморфов, причём аппендиксный. Потому что подобные морфотипы в позднем плиоцене распространены по всей Палеарктике (сочтите Число Зверя ![]() ): пумаподобные кошки Африки (предки гепардов), Азии (предки Abacinonyx и Sivapanthera), Северной Америки (предки мирацинониксов и ягуарунди). И, наконец, современная Puma concolor.
): пумаподобные кошки Африки (предки гепардов), Азии (предки Abacinonyx и Sivapanthera), Северной Америки (предки мирацинониксов и ягуарунди). И, наконец, современная Puma concolor.
Братцы, тут «чёрных» филогенетических дырок, при освещении которых можно пролить свет на филогению – хоть попой кушай:
1. Можно ли применять родовое именование Viretailurus для пумаподобных кошек из Африки и Азии, которые имеют явные морфологические отличия от европейской формы? Думаю, - нет.
Потому что есть базальные африканские позднеплиоценовые «пумы», от которых эволюционировал гепард, Acinonyx; также гипотетически имеются предковые азиатские «морфы» для более прогрессивных и специализированных других гепардообразных фелид - европейского Abacinonyx, азиатской Sivapanthera и, возможно, всей неарктической линии? Да, это пумаподобные коты, но ведь вовсе не виретайлуры.
2. Может, Viretailur-подобные кошки представляет собой некую общую циркумпалеарктическую форму пумаподобной фелины, чей ареал зоогеографически охватил всю южную область Палеарктики до 50-й параллели? Вот только в моём восприятии сам виретайлур – исключительно европеец, а схожие с ним плиоценовые кошки из Африки и восточной Азии – другие роды.
3. Не является ли Puma lacustris, как плиоценовый мигрант из северо-восточной Азии, предком вообще всех американских пумаподобных котов – мирациноникса, ягуарунди и, собственно, Puma? Если это так, то эту форму следует считать новым родом.
Не думаю, что кардинально ошибаюсь.
Р. S. По специфическим евразийским гепардам (которые, конечно же, отнюдь и совсем не гепарды) позже.
И какие же всё дохлые кошки интересные…
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#606 05 August 2024 14:39:28
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Уважаемые коллеги, только сейчас скрупулёзно добрался до Acinonyx kurteni из плиоцена Китая и выяснилось очень даже забавное обстоятельство.
В 2009 г. Кристиансен и Мазак сообщили о новом виде гепарда, основанном на черепе из позднеплиоценовых лессовых отложений бассейна Линься в провинции Ганьсу, обозвав его в честь соотечественника Бьорна Куртена как Acinonyx kurteni ( Christiansen, Mazák 2009. A primitive Late Pliocene cheetah, and evolution of the cheetah lineage), статья в свободном доступе имеется.
Голотип A. kurteni
Стоит отметить, что ничего парадоксального в этом нет, т. к. гепардоподобные кошки в китайском плиоцене присутствуют (Sivapanthera). Парадоксально другое – сочетание примитивных признаков наряду с настолько продвинутыми, которых у гепардоподобных котов в то время просто не могло быть. Это всё равно, что обнаружить в плиоценовых отложениях амазонской сельвы кости Хомо.
А потом за дело взялся Тао Денг – дядька, который в течение многих лет изучал позднекайнозойских млекопитающих именно в данном регионе Китая и на хищниках не одну собаку и кошку скушал. Ебстесснно, он испытывал неподдельный интерес к данному материалу и, тщательно пальпировав голотип Acinonyx kurteni: № SHNM8.1.07 из Шанхайского музея науки и техники, к собственному разочарованию, обнаружил, что этот образец – фальсификация (Deng, 2011. Acinonyx kurteni based on a fossil composite) – работа также есть в свободном доступе.
Зубная область краниодентальной «»мозаики», скорее всего, от сивапантеры; верхняя часть черепа – от современного гепарда, причём мозговая капсула с теменной областью – банальный гипс, а скуловые дуги – вообще не кошачьи. Такая массивность скул и Вашего покорного слугу всегда удивляла, потому что даже при множестве постмортальных факторов в процессе фоссилизации не может вызвать подобное – это даже не деформация, а сверхъествественная остеопатология. Это ж надо было так "размазать"... Да сами взгляните на фото типового экземпляра.
И как мне всё же нравится товарищ Денг, аргументировано и корректно вставившем перо в задницу европейским буржуям. Читать его «разгромную» статью – одно удовольствие. Вот, чесслово, братцы, - насколько весомо и, в то же время, «обтекаемо», ставя под сомнение лишь вероятность того, что они лично не изучали образец (скорее всего, только по фото) и выражая сожаление, что подобным актом они дискредитируют компетентность палеонтологов как учёных. Сами почитайте статью, а я Тао Денгу аплодирую даже ступнями ног. Он восхитительный и клёвый чувак (да простит он мне подобную фамильярность)!
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#607 06 October 2024 17:24:37
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Давно хочется восстановить таксономическую несправедливость в отношении евразийских пумаподобных кошек.
Первую находку описал ещё Оуэн (Owen, 1846) на основании m1 - Felis pardoides, указав на крупного хищника, размером и строением схожего с леопардом. Затем Вирэ (Viret, 1954), обнаружив в Сен-Валье череп и нижнюю челюсть, обозначил находку как Panthera schaubi. Позднее Хэммер (Hemmer, 1965) отнёс этот экземпляр к новому роду Viretailurus на основе отличий от пантер, этого же мнения придерживался и Куртен (Kurtén, 1976), также указав, что к этой форме мог относиться зуб, описанный Оуэном. Но первой, кто позиционировал эту кошку как евразийскую эволюционную линию современных пум (Felis (Puma) sp.) была Сотникова (1976). В итоге Хэммер (Hemmer, 2001), описывая эпивиллафранкские остатки из Унтермассфельда, указал, что все евразийские находки пумаморфов следует отнести к новой комбинации Puma pardoides, поскольку родовое название Viretailurus и видовое название schaubi являются младшими синонимами Puma и pardoides соответственно.
Коллеги, но ведь находки Viretailurus известны с 3 млн (Квабеби), тогда как самые ранние образцы пумы как таковой (Puma concolor) датируются лишь 0.7 млн. И если сейчас модно зачислять ягуарунди (Herpailurus) в род Puma на основании только анализа генетических данных без сколько-нибудь вменяемого морфологического сравнения, то больше того - в эту авоську на уровне подрода уже включили и Miracinonyx (Cherin et al., 2013; Chimento et al., 2014, Hemmer, 2022).
Тогда вызывает справедливое недоумение: а почему вы ещё гепарда сюда не приплели? Ведь предками Acinonyx тоже были некие пумаморфные коты… Вангую, что подобное бесовство не за горами.
А ведь громаднейший хроногеографический хиатус между пумаподобными фелинами Голарктики и Неарктики налицо. И почему не объединить эти близкородственные формы в единую группу типа трибы? Ан нет, лучше всё нивелировать и запутать (потому что филогенетический анализ всегда очень поверхностный).
Ниже предлагаю приватный морфоанализ европейских и американских образцов. Безусловно, сходство налицо, однако это подтверждает не столько принадлежность к общей группе, сколько присутствие у евразийских пумаморфных кошек архаичных признаков, свидетельствующих о меньшей специализации.
Единственный известный в настоящее время полный череп (QSV 136) найден в средневиллафранкском местонахождении Сен-Валье (Saint-Vallier) и поэтому ограничусь дентогнатальными признаками. Для верхних зубов наиболее отличительным и диагностическим в группе пумагепардовых кошек является Р4: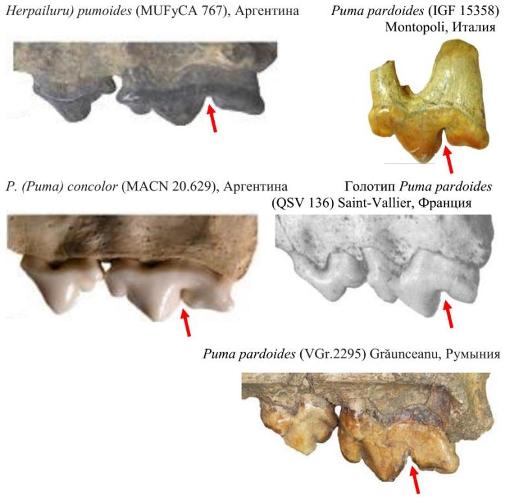
P4 Viretailurus явно отличается от Puma concolor. Протокон позиционируется более кпереди, на одном уровне с парастилем (несколько постериорно у Puma), но менее выражен (хорошо выражен у Puma ); углубление между параконом и метаконом чётко обозначено, тогда как у Puma дистальная грань паракона и метакон почти образуют слитное лезвие. Для примера приведён фрагмент с Р4 плиоценовой Herpailurus pumoides, что подтверждает ранний морфотип этого зуба у евразийских «пум».
Нижних челюстей известно больше, разной степени сохранности и датировки. К примеру, образец из французского типового местонахождения Сен-Валлье датируется 1.8 млн, из германского Унтермассфельда - 1,07 млн; и это свидетельствует, что специфические признаки у виретайлуров были неизменными. В выборке использованы несколько экземпляров, чтобы минимизировать вероятность индивидуальных изменений; также включена мандибула ягуарунди, иллюстрирующая плезиоморфность некоторых признаков.
Отличия наблюдаются следующие:
1. Симфиз у Viretailurus менее глубокий (как и у Herpailurus).
2. Венечный отросток дорсально более широкий (как и у Herpailurus), чаще заострённый у Puma.
3. Срединный гребень массетерной ямки, ответвляющийся от переднего гребня венечного отростка, хорошо выражен (рудиментарен у Herpailurus и Puma).
4. Массетерная ямка достигает задней границы m1 (как и у Herpailurus), у Puma она продлена до середины этого зуба.
5. Нижнечелюстная вырезка у Viretailurus более глубокая.
6. Мыщелковый отросток у Viretailurus более развит.
7. Угловой отросток у Viretailurus (как и у Herpailurus) менее развит и не отклонён книзу, образуя одну линию с вентральной границей горизонтальной ветви.
8. Передний бугорок на р4 (парастилид) у Viretailurus более развит.
9. Дистальный допбугорок на р4 (метастилид) у Viretailurus более развит.
10. Параконид на m1 у Viretailurus передне-задне крупнее, у Puma более заострённый.
11. Расстояние между m1 и поднятием передней границы короноидного отростка у Viretailurus больше, круто поднимается у Puma.
В итоге приватное мнение заключается в том, что родовое именование Viretailurus следует всё же сохранить, осознавая, что для подтверждения или опровержения валидности этой формы необходимы дальнейшие находки.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#608 04 November 2024 18:14:45
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Коллеги, некоторые приватные соображения по крупным кошкам из Сивалика. В частности, по Panthera dhokpathanensis и Sivapardus.
Обе формы были описаны Бакром (Bakr, 1969. «A new genus of large cat from Upper Siwaliks». Pakistan Journal of Zoology. 1 (2): 135–140) и номинально подтверждаются (Ghaffar et al., 2009. «Predator-prey relationship (Cervidae & Carnivora) and its impact n fossil preservation from the Siwaliks of Pakistan». The Journal of Animal & Plant Sciences 19(1): 54-57).
1. Panthera dhokpathanensis Bakr 1969 – голотип P.U.P.C. № 83/3, левая ветвь нижней челюсти. Ещё одна находка: P.U.P.C. № 2002/02, также левая горизонтальная ветвь мандибулы (Ghaffar, Akhtar 2005).
Называть этого хищника пантерой выглядит самонадеянно потому, что в это время настоящих пантер ещё не было, хотя размер и соответствовал мелкоразмерной самке современного леопарда. Но тут есть следующие причины: эволюционные изменения фелид в сравнении и экстраполяции на другие таксоны.
Датировка P. dhokpathanensis: Dhok-Patan/понт/поздний миоцен/7.2-5.3 млн. И тут самое интересное: полный набор зубов у P. dhokpathanensis: р1-m2. Такое присуще только проайлурам. У псевдэйлуров терминальные зубы в нижнечелюстном ряде (р1 и m2) редуцируются или вообще отсутствуют. Но у Psedaelurinae подобные признаки встречаются только у архаичных таксонов, датируемых около 10 млн. У прогрессивных форм раннего/начала среднего миоцена даже р2 уже редуцирован и однокорневой (2 корня у «P. dhokpathanensis»). Кроме того, у других сиваликских псевдэйлуров и более раннего возраста (Sivaelurus, 13,8 млн), р1 и m2 также уже отсутствуют.
Так что предварительный вывод заключается в том, что образцы Panthera dhokpathanensis принадлежит вовсе не кошке, а крупным образцам Promellivora/Eomellivora. Тем более, что размерно вписываются, да и по морфологии очень схожи.
2. Sivapardus Bakr 1969 – голотип U.Z. No. 67/22, правая нижнечелюстная ветвь. Единственный образец. Описан как принадлежащий крупной гепардоподобной кошке (крупнее леопарда) из верхнего Сивалика.
Единственное отличие от сосуществовавших Sivapanthera (brachygnathus) : в массетерной ямке нижней челюсти, которая у гепардоморфных кошек типа pardinensis и Sivapanthera простиралась до заднего конца m1, тогда как у Sivapardus она резко заканчивалась задолго до m1 и конец ямки был четко выраженным и глубоким.
Братцы, образец только один. И, во-первых: данный признак без выборки вряд ли может выглядеть диагностическим, т. к. эта черта вполне может объясняться индивидуальной или онтогенетической особенностью. Во-вторых: нелогично и сомнительно, что в одно и то же время жили экологически специфические и соразмерно одинаковые высшие хищники.
Так что это - Sivapanthera.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#609 30 December 2024 18:10:26
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Дорогие и уважаемые братцы и коллеги!
В преддверии Новогодья высылаю ссылку на архив, в котором собраны почти все статьи по ископаемым кошкам за минувший год. И в этом мне искренне помогли А. В. Лавров (ПИН), А. В Шишигин (ПГГПУ), а также Х. Мадурелл-Малапейра (Испания) и В. Семионовски (США).
https://transfiles.ru/zz7uu
Содержание следующее:
Argant 2024. A history of research on the cave lion Panthera spelaea.
История палеонтологических исследований пещерного льва.
Argant, Argant. 2024. About the origin and environment of Panthera spelaea.
Рассматривается вопрос, вопрос о том, могла ли Panthera gombaszogensis эволюционировать в P. spelaea.
Barrett,Hopkins 2024. Mosaic-evolution-underlies-feliform-morphological-disparity.
Исследование эволюции морфологического разнообразия кошачьих , основанное на данных по современным и ископаемым таксонам.
Chatar et al., 2024. Growing sabers: Mandibular shape and biomechanical performance trajectories during the ontogeny of Smilodon fatalis.
Доказательства, указывающие на то, что Smilodon fatalis претерпел изменения формы нижней челюсти на более поздних этапах онтогенеза, чем современные львы, также утверждается, что молодые особи S. fatalis могли оставаться под родительской опекой дольше, чем львы.
Deutsch et al., 2024. Myological and osteological approaches to gape and bite force reconstruction in Smilodon fatalis.
Исследование силы укуса Smilodon, дающее оценки, аналогичные таковым у ягуара.
Gimranov et al., 2024. Cheetah Acinonyx pardinensis (Felidae, Carnivora) from the Early Pleistocene of Crimea (Taurida Cave).
Описание первого ископаемого материала Acinonyx pardinensis из Крыма.
Hemmer 2024. Тhe evolution of the palaeopantherine cats, Palaeopanthera gen. nov.
Череп крупной кошки, первоначально описанный как Panthera blytheae и интерпретированный как сестринский вид P. uncia позиционируется как новый род Palaeopanthera.
Jiangzuo et al., 2024. Massive early Middle Pleistocene cheetah from eastern Asia shed light onto the evolution of Acinonyx in Eurasia.
Описан ископаемый материал Acinonyx pleistocaenicus из среднего плейстоцена Китая, представляющего самого позднего и самого крупного представителя вида; авторы считают A. pleistocaenicus видом, отличным от A. pardinensis и интерпретируют A. intermedius как мигрировавший из Африки в Азию около границы раннего и среднего плейстоцена и заменивший A. pleistocaenicus.
Lopatin et al., 2024. Mummy of a juvenile sabretoothed cat Homotherium latidens.
Описание мумии котёнка Homotherium latidens из Якутии.
Manzuetti et al., 2024. Body mass of a large-sized Homotheriini (Felidae, Machairodontinae) from the Late Pliocene-Middle Pleistocene in Southern Uruguay.
Приведены оценки массы Xenosmilus из Уругвая, представляющей собой крупнейшего гомотерина в Южной Америке начала среднего плейстоцена и одного из крупнейших саблезубов, известных на сегодняшний день.
Puzachenko et al., 2024. Panthera_spelaea_Distribution_history.
Обзор распространения Panthera spelaea в Евразии и Северной Америке на протяжении всей её эволюционной истории.
Puzachenko, Baryshnikov 2024. Panthera_spelaea_Geographical temporal variability.
Анализ изменчивости размеров нижней челюсти пещерного льва, включая половой диморфизм.
Sabol, Puzachenko 2024. Distribution of cave lions (Panthera spelaea ssp.) in mountain areas.
Распространение пещерных львов в горных районах на протяжении всей эволюционной истории.
Salesa et al., 2024. New insights on the ecology and behavior of Machairodus aphanistus.
Доказательства того, что образцы Machairodus aphanistus из миоцена Испании были поражены дентальными патологиями, которые снизили их охотничьи способности, и интерпретируют временное выживание изученных особей, несмотря на патологии, как согласующееся с существованием определенной степени социальности.
Sherani & Sherani 2024. Did Panthera gombaszogensis reach the Sunda shelf.
Интерпретация нижней челюсти из Кедунг Брубус (Ява) как свидетельство присутствия Panthera gombaszoegensis в среднем плейстоцене Зондского архипелага.
Stimpson 2024. Siwalik sabrecats review and revised diagnosis of Megantereon fossils from the foothills of the Himalaya.
Пересмотр ископаемого материала Megantereonиз Сивалика и подтверждение статуса M. falconeri как отдельного вида.
Tseng 2024. Bending performance changes during prolonged canine eruption in saber-toothed carnivores: A case study of Smilodon fatalis.
Исследование нюансов прорезывания взрослых клыков у Smilodon.
Tura-Poch et al., 2024. The disappearance of European dirk-toothed cats.
Описание фрагментарного нейрокраниума Megantereon из конца раннего плейстоцена Испании, представляющего собой самую последнюю находку Megantereon в Западной Европе, и интерпретация вымирания европейского мегантереона как связанное с климатическими изменениями.
Други мои и, надеюсь, соратники и единомышленники, с Новым Годом! Всем мой поклон, улыбка и крепкое рукопожатие.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#610 26 March 2025 21:59:19
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Коллеги, а ведь эволюция мелких кошек (FELINAE) тоже очень интересна и многогранна. Даже с вычетом таких харизматичных представителей этого подсемейства, как рыси, гепарды и пумы.
Каракаловые – это не рыси и не фелины, а отдельная группа, палеолетопись которых явно начинается 4 млн, Но есть находки рысеподобных кошек из центральной и восточной Африки (Toros-Menalla, Awash 7, Lemudong’o), датируемые поздним миоценом, 7 млн. И это фелины, дентоморфологически отличающиеся от иссуэрских рысей. В этой группе также присутствуют сервалы и золотистые коты, aurata.
Восточные коты – Prionailurus и манул, Otocolobus. Эндемики Азии. Фенотипических вариаций – хоть попой кушай. Ископаемые морфы отличительны также размерно от современных и известны из раннего-среднего плейстоцена (1.3–0.7 млн) Китая и Индонезии.
Американские коты – эндемичная неарктическая группа, включающая оцелотов, маргаев, онцилл и пр. По сути, очень коэволюционная, потому что все представители морфологически очень идентичны, а поэтому, эволюционно сравнительно молодая. Все диагнозы по таксономической принадлежности исключительными не являются и при соразмерной морфометрии по одному признаку идентифицировать виды затруднительно. В то же время это, разумеется, монофилетическая группа, судя по увеличению и топологии энтотимпанальной части слуховой буллы, форме пресфеноида, посторбитальных отростков и резцовой области, наличию Р2 и пр. Официально род Leopardus фиксируется со среднего плейстоцена (и в Сев. Америке), но есть личное мнение, что миоценовая “Felis”/Lynx longignathus из Орегона – вовсе не рысеподобная кошка, а именно «провозвестник» оцелотовой линии…
Felis разделяется на афроазиатскую и евразийскую линии. Известен, как минимум, с плиоцена Африки (Kanapoi, Laetoli, Hadar) и передней Азии (4 млн), что, вкупе с фиксацией Felis bieti свидетельствует об азиатском генезисе группы.
Азиатские коты – также эндемичные кошки (пардофелисы и катопумы), краниодентально и морфологически наиболее базальные из «продвинутых» фелин типа рысей и крупноразмерных пумо/гепардообразных, но по архитектуре черепа отмечается явная схожесть с архаичными пантеринами (Neofelis, дымчатый леопард). В палеолетописи фиксируются со среднего плейстоцена Китая (Hualong, Bailongdong), но филогенетически выглядит сестринской по отношению как к рысеподобным, так и к линии Acinonyx/Puma и Pantherinaе.
И есть ещё очень интересная группа позднемиоценовых пристифелисовых кошек (7 млн). В настоящее время официально валиден только Pristifelis attica, однако подобные коты известны не только из Европы, а также из Передней Азии, Северной Африки, Китая и даже из СевАмерики (?). Так что там, более чем вероятно, несколько родов. А на самом деле, это скорее всего, целая триба.
В каждой из этих линий были как «аппендиксные», так и прогрессивные векторы. И история Felinae на самом деле не менее многовластна и жизнелюбива, чем саблезубых кошек и пантерин.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#611 27 March 2025 14:08:06
- Roman Shevchenko
- Любитель животных
- Зарегистрирован: 14 February 2021
- Сообщений: 720
Re: Эволюция кошкообразных (Feliformia)
А среди пантер были генералисты весом в 5-7 кг.? Или это подсемейство изначально шло в сторону укрупнения размеров, то есть было ориентировано на крупную добычу?
И мне кажется, что мелкие кошки на деле самые архаичные именно как подсемейство. Не могу точно сказать, но именно среди них сохранились довольно генерализированные мелкие формы. Может, именно от более архаичных представителей этого подсемейства пошли пантеры и саблезубые кошек? Опять же лишь предполагаю.
Просто я не думаю, что домашняя кошка в каких-нибудь в морфологических признаках прогрессивные льва.
Неактивен
#612 29 March 2025 00:09:12
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Роман, спасибо за реакцию, а то я в этих кошковетках в последнее время сам с собой общаюсь и, чессгря, поднадоело…
В приватном воззрении понятие «прогрессивности» применимо лишь при морфологическом анализе всей группы, причём с учётом ископаемвх остатков. К примеру, если взять линию Acinonyx из пумагепардовых кошек по зубам верхнего ряда, то у раннеплейстоценовых гепардов (Swartkrans, Makapansgat, Koobi Fora - 1.9-1.8 млн) P3 крупный, с высоким главным бугорком и с массивными дополнительными вершинками. Этот зуб отличается от поздних форм тем, что передняя вершинка не находится на одной линии с главным бугорком, а его задняя часть уширена. И этим он очень сходен с плиоценовыми образцами (Laetoli, Woranso-Mille, Awash 7, Omo Shungura C-Usno 3.6-3.0 млн), которые на самом деле, скорее всего, ещё не гепарды, а африканские пумаподобные кошки (вангую, что новый таксон). Затем, уже заметна разница в положении протокона на P4: у среднеплейстоценовых гепардов расположен более антериорно, тогда как у позднеплейстоценовых и современных протокон на Р4 редуцирован и это является диагностичным признаком всех гепардов. Также его положение чётко отличает Acinonyx от Miracinonyx (американских «гепардов»).
Здесь налицо прогрессивность. Но, в то же время, это понятие связано со специализацией. И, если в общем плане, прогрессивность (апоморфия/синапоморфия) у кошачьих выражается в гипертрофии верхних клыков (режущая функция у махайродонтных и давяще-удушающая у фелин/пантерин), дентальной сепарации в карнассиальных зубах (исчезновение первых/последних премоляров/моляров), редукции зубов/бугорков, не участвующих в окклюзии; как и связанной с ней черепной архитектурой (начиная от лицевого профиля и заканчивая размером и позицией мастоидных отростков, потому что всё это согласуется по характеру, динамике и алгоритмам нанесения первых, летальных укусов и последующего акта умерщвления). А по прогрессивности – все живущие в нынешнее время кошки таковые, потому что выжили в эволюционных сражениях на протяжении миллионолетий. Гепарды, рыси, львы и наши Васьки с Машками.
Но с точки зрения фелидного морфотипа самый и наиболее кошачий, апогей и высшая точка – манул, Otocolobus.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#613 29 March 2025 00:10:19
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Теперь по генезису Pantherinae. То, что фелиды сменили Nimravidae, связано с с начавшимся планетарным похолоданием и другими климатическими изменениями в раннем миоцене, потому что происходила замена лесных ассоциаций на лесостепные, возникли степи и саванны, а подобные изменения вели к увеличению ландшафтной мозаичности и появлению свободного экологического пространства. Эти ниши занимали новые группы травоедов, а их более сложные поведенческие реакции и морфофизиологическая пластичность позволяли кошкам настолько успешно конкурировать с экологически сходными нимравидами, что эти примитивные фелоиды к концу среднего миоцена почти полностью исчезли.
Пантерины, очевидно, действительно, изначально экологически специализировались на доступных крупноразмерных жертвах. Возможно, локализация их охотничьих угодий включала прибрежные зоны водопоев копытных в менее залесённых участках местности. Меньшие размеры в сравнении с амфиционами, перкрокутами и урсидами позволяли архаичным пантерам успешней скрадывать добычу, а массивный череп с выраженным сагиттальным гребнем, широкими скуловыми дугами и челюсти с крупными зубами – атаковать и довольно быстро умерщвлять относительно крупную добычу типа ювенильных или ослабленных особей. И хотя по уровню специализации зубного аппарата они выглядели уже специализированными миофагами, в экологическом отношении они, пожалуй, вряд ли очень отличались от современных кошек Юго-Восточной Азии и рысеподобных фелид.
Генерализированная форма пантеровых, известная в настоящее время - Neofelis, дымчатый леопард. Фоссилии фиксируются с раннего плейстоцена, 1.2 – 0.9 млн.
Интересно, что в «палеопланктоне» подсемейства Psedaelurinae имеется группа, вполне претендующая на место протопантер, занимавшая в раннем-среднем миоцене ареал от Испании до Китая и, будучи крупнее Styriofelis, в размерном диапазоне от рыси до мелкого леопарда (Miophantera, Sivaelurus, Vishnufelis). Но это ещё – псевдэйлуры, у них ещё есть р2, алисфеноидный канал, широкий M1 и ряд других архаичных признаков, которые присутствуют и у примитивных Machairodontinae. Хронология кластера этих кошек 16 – 8 млн, тогда как Panthera blytheae (ещё под вопросом, пантера ли это) и Pachypanthera датируются 6 млн, только эти котятки уже очень даже крупные.
Самый близкий к обобщённому (генерализированному) типу пантерин - Pseudaelurus (Schizailurus) сf. lorteti (Qiu, Gu 1986; Wang et al., 1998; Jiangzuo et al., 2020), образец V 8070 – фрагмент левой нижнечелюстной ветви из Сяцяовань (Xiacaowan). Наряду с изолированными находками р4 и m1 эта форма дентоморфологически более близка с евразийской Miophantera, чем с другими китайскими образцами, только его датировка – 16 млн. к тому же это – явный псевдэйлур.
Так что имеется некоторая темпоральная лакуна минимум в 2 млн. А это при отсутствии близлежащих адекватных фоссильных находок всё же много. Даже с точки зрения палеонтологии.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#614 29 March 2025 14:59:22
- Roman Shevchenko
- Любитель животных
- Зарегистрирован: 14 February 2021
- Сообщений: 720
Re: Эволюция кошкообразных (Feliformia)
Corvin :
Но с точки зрения фелидного морфотипа самый и наиболее кошачий, апогей и высшая точка – манул, Otocolobus.
Можете, пожалуйста, расписать про прогрессивность манула. Про его морфотип. Форуму нужны научные описания подобного плана.
Специализация больших кошек к добыче крупной дичи может считаться в чём-то прогрессивным, но это действительно больше всё же специализация.
Впрочем, кошачьи-изначально специализированное к западной охоте семейство гиперплотоядных зверей(даже самые первые и архаичными уже таковыми были, судя по останкам). Потому они и оказались столь успешными.
Отредактировано Roman Shevchenko (29 March 2025 14:59:47)
Неактивен
#615 29 March 2025 15:05:02
- Roman Shevchenko
- Любитель животных
- Зарегистрирован: 14 February 2021
- Сообщений: 720
Re: Эволюция кошкообразных (Feliformia)
Причем самое интересное, что первые кошкообразные(вивверариды и после них), а также многие виверровые и пальмовые циветы-всеядные. Но по каким-то причинам в этом плане они уступили псообразным. Может, тут дело в каких-то морфологических признаках?
Неактивен
#616 29 March 2025 15:20:52
- Roman Shevchenko
- Любитель животных
- Зарегистрирован: 14 February 2021
- Сообщений: 720
Re: Эволюция кошкообразных (Feliformia)
Есть ещё одна интересная мысль-у гиен, в отличие от псовых, долгое время существовала полигамия, которая сохранилась у пятнистых гиен. Лишь позднее она появилась у некоторых видов. А всё по одной причине-переход с моногамии на полигамию не выгоден, особенно для стайных хищников.
Отредактировано Roman Shevchenko (02 April 2025 13:14:51)
Неактивен
#617 01 April 2025 20:29:15
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Roman Shevchenko :
Можете, пожалуйста, расписать про прогрессивность манула. Про его морфотип. Форуму нужны научные описания подобного плана.
Без проблем. Дабы не быть голословным:
Верещагин Н.К. Сравнительная краниологическая характеристика диких кошек СССР //Зоологический журнал 1967, т. XLVI вып.4.
В этой работе Николай Кузьмич всё подробно и обстоятельно объясня(л)ет.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#618 09 April 2025 16:08:55
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Стараюсь разобраться, насколько возможно, в эволюции Felis… Братцы, там не менее интересно, как у пантер, саблезубых и рысеобразных, о чём всегда подозревал.
Ну, то, что наши Васьки это потомки lybica, наверное, всем давно известно. И что их степень гибридизации с silvestris выглядит довольно мизерной, наверное, тоже. Хотя, на самом деле, это вызывает удивление.
Но наиболее забавное в том, что когда в подсемействе Felinae «отсекаешь» рысей, каракаловых и пумагепардов, а также южноамериканских (оцелотов, маргаев) и восточноазиатских (Pardofelis, Catopuma) котов, то даже в этой группе масса белых пятен по эволюции, филогении и систематике. А это всего лишь палеарктический Felis, из которых фенотипически и размерно выделяются разве что хаус и китайский bieti. И палеолетопись Felis хоть и наиболее богата, конечно же, начиная с остатков в археологических памятниках мезо/неолита, но это уже доместикационная ипостась. Однако, на самом деле, эта линия эволюционировала, как минимум, с конца плиоцена (2.5 млн).
Из ныне живущих форм валидны Felis lybica (степной кот), F. margarita (барханный), F. nigripes (чернопалый), F. chaus (камышовый), F. silvestris (лесной), F. bieti (китайский) и ещё выделяют F. catus – домашнюю кошку. Большинство из них известны со среднего плейстоцена (0.8-0.7 млн) и, ввиду некоторой робустности отдельных краниомандибулярных позиций, как и архаичном укрупнении премоляров (Р3/р3) в сравнении со современными, позднеплейстоценовыми, их даже можно считать хроноподвидами. Но вот «фелисы» раннего плейстоцена Африки, Азии и Европы (1.9 млн) - явно более генерализованные формы, которые достойны выделения в отдельные виды. Здесь - поле непаханное.
И это не миоценовые Pristifelis. Пристифелисовые – это группа фелид, вообще относящихся к отдельной архаичной трибе.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#619 08 May 2025 17:10:44
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Коллеги, в опровержение поста некоего хомячкового псевдоспециалиста по фелинам, нахватавшегося «верхушек», но вряд ли читавшего оригинальные статьи (случайно наткнулся и весьма возмутило):
https://vk.com/wall-158490065_314071
Впервые Felis minuta упомянута в архаичной работе Шмерлинга (Schmerling, 1833). В 70-х гг. ХХ века она упоминается (несмотря на заверения автора) лишь фрагментарно (Friant 1970, Clot 1975). Даже Бьорн КуртЭн (Kurten), издавший в 60-х гг обстоятельные монографии по эволюции и морфологии европейских и ближневосточных фелисов, о данном таксоне вообще не упоминает. А этот дядька – крутой и авторитетный, его работы даже полувековой давности с карандашом читать следует.
Братцы, Felis minuta – это таксон nomen dubium, поэтому в официальных публикациях он встречается редко. Голотип Шмерлинга вообще основан на двух плечевых костях, причём разновозрастных. Посему – сами понимаете… Все остальные обращения основываются лишь на мелкоразмерности образцов в сравнении с такими же близлежащими фоссилиями «сильвестриса».
Почему никто не обращает внимания на онтогенез? Если сравнивать черепа Felis silvestris молодого и матёрого самца и самок таких же возрастов – это будут несколько различные черепа четырёх кошек. Причём если не знать возрастные и половые изменения краниума – это вообще в воззрении некомпетентных специалистов 4 разных вида будут)).
К сожалению, работ 2012-20 гг., на которые ссылается автор, не нашёл, а вот упоминания о находке костей фелиса в неандертальской стоянке Мула-Герси (Baume Moula-Guercy) обнаружил. Но – лишь в фаунистических списках.
Так что, такой кошки, как F. minuta не существует. Это – подростковые особи позднеплейстоценовых (120-90 тлн) F. silvestris, у которых молочные зубы уже сменились постоянными, но нижнечелюстная ветвь ещё не оформилась в векторе утолщения (робустности).
На самом деле странно ожидать, что в европейских биомах сосуществовали две формы экологически идентичных котов. Такого не бывает. Или очень кратковременно, не оставляя следов в палеолетописи.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#620 11 June 2025 15:49:29
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Братцы, а я всё же чуть-чуть разобрался в «фелинной» миоцен-плиоценовой каше ЕвроАфрики…
Представители Felinae появляются в этих регионах в конце миоцена. Генезис –разумеется, некие продвинутые псевдэйлуры и, очевидно, из Сиваликов.
Первичные – это миоценовые фелины из Европы (Pristifelis), Африки и Китая. Все эти кошки примечательны двукорневым Р2 (у Felis он однокорневой), количеством молочных зубов и также отличаются от Lynx отсутствием р2 (который у рыси встречается в 20% образцов) + укороченная постклыковая нижнечелюстная диастема. Отличия заметны но, к сожалению, ограничены почти только дентомандибулярным материалом и ограниченным посткраниальным. В целом, эти архаичные кошки (размером со среднеразмерных рысей) таксономически можно распределить следующим образом:
1. Миоцен-плиоценовые пристифелисы Европы (Pristifelis): ранняя форма attica (MN 12, средний туролий) и поздняя сhristoli (MN 13-14, поздний туролий-ранний валлезий) в ряде работ (Rook et al., 1991; Sardella, 2008; Bartolini-Lucenti et al., 2021) объединяются в единый хроновид практически только на основании разницы в размерах – сhristoli крупнее. Однако морфология свидетельствует не только о сходстве таксонов (один род), но и заметна разница (у сhristoli более спрямлённая вентральная граница нижней челюсти, менее высокий венечный отросток, больше высота между угловым и мыщелковым отростками; более редуцирован р3 и укрупнённая группа р4-m1 и пр. признаки, свидетельствующие о явной прогрессивности). Поэтому знак тождественности как единого вида ставить, наверное, преждевременно. 
Слева: голотип attica BSPM – РIК A 511.116, Пикерми (Bayerische Staatssammlung fur Palaontologie und Geologie) г. Мюнхен, Германия Справа: голотип сhristoli, музей естественной истории Базеля (Natural History Museum Basel), Германия. Монпелье (Montpellier).
2. Миоценовые фелины Африки (7–5.5 млн). Материал из Чад (Toros-Menalla) и Кении (Lukeino, Lemudong’o) свидетельствует о присутствии кошки, морфологически и размерно находящейся в диапазоне Pristifelis и, очевидно, что все образцы принадлежат к едииой архаичной морфогруппе (Barry 1987, Geraads 1997; Werdelin, Dehghani 2010; Werdelin, Manthi 2012; Raynal, Mohib 2016). Вид это или отдельный род предвещать сомнительно, но однозначно материал относится к группе пристифелисовых котов. Возможно, от этой линии позже, в плиоцене, эволюционировали каракаловые.
3. Миоценовые фелины из восточной Азии (Китай и Монголия). Датировка 8.0-5.5 млн. Находки из Люфенг, Шаньси и пр. (Zdansky, 1924; Qi, 1983; Qiu, Qiu 1995; Roussiakis 2002) Сходны с Pristifelis морфологически, но отличаются меньшим размером, укрупнённым клыком, развитым p3: более остроконечным, со спрямлённой позицией (наклонной к р4 у Pristifelis) и более развитой передней вершинкой; также кулисным захождением постериорной границы р4 за параконид m1 (прямолинейное расположение этих зубов у Pristifelis).
Архетипичные образцы из Китая (Zdansky, 1924). Сверху: Shansi. loc 32; снизу: Shansi lok. 49.
Коллеги, с точки зрения хроностратиграфического контекста это гомологичный морфотип, однако дентальная морфология несколько отличительна (степень развития р3, окклюзионная позиция р4/m1). Также образец из lok. 32 (8.0 млн) отличается от lok. 49 (7.0 млн) что неудивительно, всё же их почти «лям» лет разделяет не только визуально – в промерах и пропорциональных сравнениях наблюдаются характеристики, сходные с таковыми в группе attica/сhristoli: как минимум, увеличение в размерах у более поздних форм.
Несколько сомнителен образец из loc 32, очень схожий с метайлуровыми котами. Сохранность р4-m1 не позволяет провести обстоятельный морфоанализ, однако метрически он гораздо меньше. К тому же подобный экстерьер (утолщённость горизонтальной нижнечелюстной ветви и спрямлённость симфиза) характерен не только для «пристифелисовых» - это конвергентно наблюдается и у рысеподобных фелид: робустность у плезиоморфных форм плиоцена и грацильность у плейстоценовых.
С учётом вышеизложенного смею гипотезировать, что позднемиоценовых котов восточной Азии с учётом морфологических особенностей, морфометрики и географического положения следует суммировать в отдельную форму родового ранга. Помимо прочего, эта форма потенциально может являться предковой формой для производных азиатских Prionailurus, Pardofelis/Catopuma…
Также «вангую», что со временем (по нахождению более полного материала) вышеупомянутые африканские и азиатские образцы ранжируют как отдельные роды, демонстрирующих наиболее плезиоморфные состояния дентальной морфологии среди архаичных представителей Felinae. А всю группу обозначат как эволюционную линию, происходящей от некоей кошки псевдэйлуринной категории со специализациями, параллельными таковым у Metailurus. Очень надеюсь, что как отдельную архаичную трибу Pristifeliini.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#621 01 September 2025 18:37:55
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Коллеги, уж позвольте «выплеснуться»…
Семейство Felidae – мир совершенных хищников, на протяжении последних 15 млн занимающих доминантные трофические ниши во всех биотопах, к которым они сумели адаптироваться. И современные представители кошачьих это лишь верхушка айсберга, потому что составляют не более 1/3 того эволюционного разнообразия, которое победно маршировало из среднемиоценовой древности.
В сообщениях об ископаемых котах, как правило, львиная доля))) интереса обывателей касается наиболее харизматичных представителей – саблезубых, пантерин или крупноразмерных кошек типа пумагепардовых. И это несколько забавно.
Потому что зачастую обходится стороной подсемейство Psedaelurinae, хотя именно в этой базальной группе и намечаются эмбриональные формы махайродов, смилодонов, рысей, котиков и пантер. К тому же они размерно варьировались от мелких до габаритов крупного леопарда. И присутствовали интересные «аппендиксные» формы, которые впоследствии вымерли без потомства (например, африканские Diamantofelis), едва преодолев плиоценовую границу. Причём такая же картина наблюдалась и в Неарктике, демонстрируя ряд эндемичных псевдэйлуровых, отличающихся от палеарктических, но морфологически схожих (Nimravides). Или непохожих (Pratifelis). Или не просто идентичных, а даже более прогрессивных, чем евразийские аналоги (Hyperailurictis stouti ).
Или Machairodontinae. Братцы, это – «винегретная» группа, включающая конвергентные таксоны саблезубых кошек Афроевразии (Machairodus, Amphimachairodus) и Северной Америки (Nimravides, Smilodon), но с позиций изначального генезиса, дальнейшего прохореза и морфологии базальных морфотипов они должны ранжироваться в отдельные подсемейства. Как и Metailurinae – это не фелины и не саблезубые, а самобытная группа кошачьих, эволюционировавшая параллельно вышеупомянутых. В которой экологически присутствовали эквиваленты современных фелисов и рысей (Metailurus), гепардов (Yoshi) и пантер (Therailurus/Dinofelis). К тому же морфологически (краниальная «архитектура», зубы, уши, базикраниаль) занимают срединное положение между Felinae и Machairodontinae. Но это уже другая тема и весьма специфическая.
Ну, да и Дарвин с ними… Давно интересуюсь эволюцией подсемейства Felinae. Классически очень удобно классифицировать их по линеевской систематике, т.к. трибы/подтрибы вполне наглядно отражают филогению:
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#622 06 November 2025 19:43:18
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
К эволюции рысей.
В настоящее время принято, что к роду Lynx относятся виды shansius Teilhard, Leroy, 1945; issiodorensis Croizet, Jobert, 1828; pardinus Temminck, 1827; hei Jianzuo et al., 2022; rufus Schreber, 1777; lynx Linnaeus, 1758; canadensis Kerr, 1792.
Взгляд очень консервативный, потому что шаньсийская форма это ещё не совсем рысь, а только рысеподобная кошка (об этом кулуарно чуть ли не полвека дискутируют). Так же, как и североамериканские «рыси» proterolyncis Savage, 1941; longignathus Shotwell, 1956 и rexroadensis Stephens, 1959 – вполне валидные таксоны, но морфологически и стратиграфически классифицируются как кошки, близкие к группе пристифелисовых.
Соблазнительно дифференцировать рысей на группы, причём материала достаточно для всяческих филогенетических гипотез:
1. Иссуэрско-парделевые рыси (от Китая до Европы) с рядом видов, из которых до настоящего времени дожила иберийская pardinus. Не исключено, что к этой группе относится и вымершая североафриканская thomasi.
2. Трансберингийская рысь: ископаемая китайская hei и североамериканская rufus. Рысь Хэя – аберрантная азиатская форма, а рыжая рысь прошла почти через миллионолетнюю трансформацию (calcaratus), прежде чем принять современный экстерьер.
3. Собственно, голарктическая рысь с двумя видами: евразийская Lynx lynx и канадская L. canadensis.
Одним из примеров можно привести верхний зубной ряд (масштаб не соблюдался):
A. L. issiodorensis, A1. HMV2012, Longdan; A2. AMNH 96-B1042, Xiazhuang; A3. AMNH 62-B756, Shouyang;
B. L. hei, HMV2010, Longdan;
C. L. rufus, C1. AMNH M14822; C2. AMNH M243108;
D. L. pardinus, D1. AMNH M169492; D2. USNM 152619;
E. L. canadensis, E1. USNM 077095; E2. USNM 212617;
F. L. lynx, F1. AMNH 41337; F2. IOZ T0139.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#623 09 November 2025 20:56:29
- Janika
- Любитель животных

- Откуда: Cамара
- Зарегистрирован: 02 July 2009
- Сообщений: 678
Re: Эволюция кошкообразных (Feliformia)
Пиренейские рыси такие мелкие потому что ближе к базальной форме или вторично обмельчали в изоляции ?
Потерялась на дороге жизни.
Неактивен
#624 10 November 2025 07:34:08
- Doc
- Любитель животных
- Зарегистрирован: 03 April 2016
- Сообщений: 469
Re: Эволюция кошкообразных (Feliformia)
Janika :
Пиренейские рыси такие мелкие потому что ближе к базальной форме или вторично обмельчали в изоляции ?
Или проявляется правило Бергмана - мелкие, потому что в более тёплых местностях?
Неактивен
#625 10 November 2025 19:36:55
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Эволюция кошкообразных (Feliformia)
Братцы, с рысями приходилось общаться… но, на удивление и, к сожалению, в коллекции ЗИН нет Lynx pardinus . В Питере имеется обширная коллекция черепов со всей России от Приморья до восточных границ; рыси из Фенноскандии и Средней Азии; даже американские rufus и canadensis присутствуют (массу фоток некогда «нащёлкал»), а вот парделевых (иберийских) не нашёл, каюсь... Так что приватное мнение о них пока основано на ископаемом материале issiodorensis , как филогенетически наиболее близкой формы, отштудированным статьям других исследователей и общении с А. В. Лавровым. «Вживую» материал по современным парделевым рысям щупать не пришлось, посему заранее не взыщите.
Морфологически тут немало нюансов (очевидно, конвергетных), т.к. по краниальным промерам и степени развития сагиттального и затылочного гребней она близка к канадской рыси. Также однотипно строение пресфеноида, межглазничной области, топография слуховых булл, взаиморасположение яремного и переднего суставного отверстий и прочих признаков. Вместе с тем своеобразие в других краниодентальных признаках выражено гораздо резче, чем у любого из видов/подвидов Lynx на всём пространстве Палеарктики. Так что это рысь, но совершенно другая, реликтовая форма. О происхождении можно ознакомиться в работах:
- Ficcarelli, Torre. 1977. Рhyletic relationships between Lynx group issiodorensis and Lynx pardina.
- Werdelin, 1981. Evolution of lynxes.
В настоящее время принято мнение, что pardinus происходит от issiodorensis , причём Лавров рассматривает её как угнетённую, педоморфную «морфу» (эктопарастиль на P4, метаконид на m1 и пр.), т. к. линия issiodorensis демонстрирует наибольшее географическое распространение в виллафранке а, впоследствии, по-видимому, была ограничена рефугиумами - возможно, только Испанией. Потому что уже в кромерских слоях (средний плейстоцен) по всей Европе все находки принадлежат уже только Lynx lynx .
Личное мнение: группа pardina , безусловно, имеет самые тесные филетические связи с плиоценовыми иссуэрскими рысями и представляет собой некий продукт эволюционного тренда, начавшегося в нижнем виллафранке и характеризующегося уменьшением размеров и приобретением более специализированных черт зубной системы. Уменьшение размеров связано, в основном, с рефугиальным ограничением (сужение ареала подтверждается такими зоогеографическими аналогами, как, например, распространение выхухолей в Испании). Хотя стоит отметить, что начиная с Сен-Валье и Оливола у issiodorensis уже наблюдается уменьшение размеров (рыси из Вальдарно вообще ненамного крупнее современных пиренейских). Возможно, эта тенденция связана с серией оледенений, сдвигающих ареал иссуэрских рысей южнее, в то время как «линксоподобные» формы оказались более пластичнее, заняв эконишу в бореальной фауне. Так что, Janika , в Вашей гипотезе в одном строю.
Правило Бергмана… Да почему бы и нет? Распространение испанской рыси ограничено широтами несколько южнее основного массива ареала Lynx в Палеарктике (примерно на уровне популяций Кавказа и Средней Азии). Посему это, как кажется, также весомый довод. Андрей, спасибо!
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен