Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#676 09 September 2016 19:59:06
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Ого, на вторую страницу перевалило..
Pachycrocuta brevirostris Gervais, 1850
Таксономическая история: Библиографическое исследование (Alba et al., 2015) показало, что первая публикация, в которой использовалось именование данного таксона, применено Жерве (1850), а Эймард был лишь автором коллекции, череп из которой описан в качестве типового образца. Таким образом, согласно Международному кодексу зоологической номенклатуры (МКЗН, 1999), именование Pachycrocuta brevirostris Aymard, 1846 не отвечает нормативным критериям и правильным именованием следует считать Pachycrocuta brevirostris Gervais, 1850.
Синонимы: Hyaena brevirostris, Pachycrocuta robusta
Местонахождения: Франция – Сартанетт (Sartanette), Верхейзель (Verchizeuil), Буи-де-Рикет (Bois-de-Riquet), Сензелье (Sainzelles), Сен-Прест (Saint Prest), Валлонэ (Vallonet), Сессагэ (Ceyssaguet); Италия – Фоггия (Foggia), Оливола (Olivola), Тассо (Tasso), Вальдарно (Valdarno), Пирро Норд (Pirro Nord), Касафрата (Casa Frata), Поджо Россо (Poggio Rosso); Испания – Фонелас (Fonelas SCC-1, Fonelas P-1), Вента Мицена (Venta Micena), Куэва Виктория (Cueva Victoria), Инкарнал (Incarcal), Понтон де-ла-Олива (Pontón de la Oliva), Кал Гвардиола (Cal Guardiola); Чехия – Странска Скала (Stranska Skala), Златый конь (Zlaty Kon); Германия - Зюссенборн (Süssenborn), Вюрцбург-Шальксберг (Würzburg-Schalksberg), Мейнинген (Meiningen), Унтермассфельд (Untermassfeld), Эрпфингер (Erpfinger); Великобритания - Локации Форест Бед (Cromer Forest Bed: Backton, Mundesley, Sidestrand, Westbury-sub-Mendip); Греция – Петралона (Petralona), Геракару (Gerakarou 1), Аполлония (Apollonia 1), Каламото (Kalamoto); Словакия – Гомбасцог (Gombaszog), Македония – Манастирек (Manastirec); Венгрия – Вертешсоллеш (Vertesszollos), Кишланг (Kislang); Молдовия – Тирасполь; Россия – Ливенцовка, Ахтанизовская (Приазовье), Тологой, Засухино (Забайкалье); Грузия – Ахалкалаки; Казахстан – Актогай; Таджикистан - Лахути 2; Монголия – Налайха (Nalaicha)
Голотип: череп с нижней челюстью (без номера), Сензелье (Sainzelles), Франция.
Время распространения: поздний виллафранк – средний плейстоцен (MN 18 - MQ 22); 1,8 - 0,5 млн лет.
Сравнительные описания: 
Голотип представляет собой череп (номер не указан) с ассоциированной нижней челюстью из раннеплейстоценового (1,3 млн) французского местонахождения Сензелье (Sainzelles), хранящийся в Национальном музее естественной истории г. Париж. Образец представляет крупную гиену с укороченной и широкой лицевой частью; P2 округлый, небольшой и слегка заострённый, имеется цингулум. P3 чрезвычайно массивный, передний бугорок крупный, главный не очень высокий, задний, с цингулумным пояском, выше. P4 очень крупный, наличествует метакон; парастиль большой, имеется дейтерокон; метакон не выше паракона; М1 снижен. Нижняя челюсть характеризуется массивностью и одним большим подбородочным отверстием, расположенным ниже р2; первые премоляры укорочены, но утолщённые; третий и четвёртый чрезвычайно массивны; m1 широкий, лишённый метаконида, талонид одно- или двухбугорковый.
Нижняя челюсть P. brevirostris (C4-CE8-149), Валлонэ (Vallonnet), Франция
Этой же зоной (MQ 20) датируются находки из Сен-Прест (Saint-Prest) и Валлонэ (Vallonnet), тогда как в Италии и Испании наличествуют более ранние. Впервые P. brevirostris появляется в Европе в итальянской локации Оливола. Примерно тем же возрастом (около 1,9 млн) датируются находки из Фонелас (Fonelas P-1) Испании. Вообще, в Испании имеется довольно обильный материал по находкам пахикрокуты. Она хорошо представлена в Вента Мицене (Venta Micena) примерно 1,6-1,5 млн лет назад, характеризуясь крупным размером, широкими и массивными премолярами, P4 с хорошо развитым, мезиально направленным протоконом, большим парастилем и среднеразмерным метастилем. Вообще, в данной южноиспанской локации Pachycrocuta регистрируется даже в слое EVT7, датируемом ок. 0,83 млн лет.
Верхнечелюстной фрагмент (VM2281) - P2 крупный, с небольшим передним бугорком и хорошо развитым цингулумом; P3 относительно широкий, также с развитым внутренним цингулумом; P4 с позиционированным наискосок крупным протоконом, достигающим высокого парастиля. Паракон также высок, метастиль увеличен.
Нижняя челюсть Pachycrocuta brevirostris (VM2278), Вента Мицена (Venta Micena), Испания
Гнатодентальный материал представлен лучше. Левые нижнечелюстные ветви (VM2278, VM2271) соответствуют очень крупному и мощному представителю семейства Hyaenidae, они шире в районе p3, чем у р4. И демонстрирует мощный жевательный аппарат с хорошо развитыми премолярами. Нижние клыки заострённые, трегуольные в основании – даже массивнее, чем у сосуществовавших представителей рода Ursus. p2 (VM2271, VM2278; VM10325) имеют высокие протокониды; внутренний цингулум развит слабо. р3 (VM2271, VM2278; VM10382) также с высоким протоконидом, средний бугорок редуцирован и спозиционирован лингвально, задний развит хорошо, чётко отделяясь от протоконида. p4 (VM2271, VM2278; VM10382; VM10322) демонстрирует высокий индекс пропорций (ширина/длина), с крупным задним бугорком, который выше, чем средний; цингулумный поясок намечен. m1 (VM2271, VM2278; VM10324; VM10322; VM2276) с хорошо развитыми протоконидом и параконидом, талонид двухбугорковый, метаконид отсутствует.
Хороший материал наличествует и в местонахождении Куэва Виктория (Cueva Victoria), хронологически относящемуся к возрасту 1,1 млн лет назад. P2 небольшой, с обособленным протоконом и развитыми средним и задним бугорками. P3 широкий, с высоким протоконом; задний бугорок небольшой, но чётко обозначен, цингулум чётко обозначен. P4 крупный, парастиль развит умеренно, вершинка метастиля снижена, имеется задний внешний цингулум, величина которого вариативна. М1 по сравнению с премолярной серией небольшой, наличествует цингулумный поясок, метакон редуцирован. Нижние зубы: p2 с высоким протоконидом; p3 с хорошо развитым протоконидом и сжат с боков; р4 также с развитым протоконидом и крупными средним и задним бугорками; m1 с хорошо развитым двухбугорковым (гипоконид и гипоконулид) талонидом.
Левая нижнечелюстная ветвь P. brevirostris (EVT 24641), Cal Guardiola (Испания)
Также более сотни образцов извлечено из эпивиллафранкских горизонтов в Унтермассфельде (Untermassfeld) с датировкой 1,07 млн лет назад – различного возраста, плюс костные остатки травоядных с погрызами и множество копролитов. 
Нижнечелюстная ветвь P. brevirostris (IQW 1980/15 918), Унтермассфельд (Untermassfeld), Германия
Также немалый интерес представляют и восточно-европейские находки раннего плейстоцена. Часть правой ветви нижней челюсти (GIN 1126/1) из Ахтанизовской (Таманский полуостров), датированный 1,1 млн лет назад, принадлежит P. brevirostris ввиду массивности нижней челюсти, крупного подбородочного отверстия под р2, массивности премоляров и морфологии р3-p4: Нижнечелюстной фрагмент P. brevirostris (GIN 1126/1), Ахтанизовская (Россия)
Нижнечелюстной фрагмент P. brevirostris (GIN 1126/1), Ахтанизовская (Россия)
Образец принадлежит взрослой особи и его морфологические признаки хорошо согласуются с европейскими образцами P. brevirostris. В то же время весьма интересен нижнечелюстной фрагмент из Ливенцовского карьера (RSU-231) хапровского комплекса: 
Нижнечелюстной фрагмент Р. brevirostris (RSU-231), Ливенцовка, (Приазовье) Россия.
Нижние резцы и клыки крупные. Второй премоляр с небольшим передним дополнительным бугорком и крупным задним. Размеры р2 превышают вариативный предел изменчивости для Р. perrieri и соответствуют размерам Р. brevirostris. Стёртый рЗ более массивный, чем р4 и обладает небольшим передним дополнительным бугорком, а также хорошо развитым переднелабиальным цингулумом. Хищнический зуб относительно р4 удлинён, метаконид отсутствует, талонид двухбугорковый. От типового экземпляра ливенцовская Pachycrocuta отличается меньшими размерами, наличием переднего дополнительного бугорка на р2 и более редуцированным р4.
В принципе, P. brevirostris из Ливенцовки близка к виллафранкским формам по таким признакам, как наличие передних бугорков на р2 и р3 и умеренно редуцированного талонида с двумя бугорками, а по степени редукции р4 она всё же сходна с P. licenti из плиоцена Китая. В целом для хапровского фаунистического комплекса характерно некоторое сходство с азиатскими аналогами средневиллафранкских фаун и по некоторым признакам Pachycrocuta из Нихэвань близка к ливенцовской гиене. Таким образом, присутствие Р. brevirostris в фауне хапровского ком¬плекса указывает на то, что проникновение Pachycrocuta в восточную часть Европы про¬изошло раньше, чем в западные регионы – то есть, в малоазиатском регионе.
По наблюдению Сотниковой (Сотникова и др., 2002), у P. brevirostris фаун конца раннего и среднего плейстоцена Европы преобладает m1 с одним бугорком на талониде, тогда как у виллафранкских форм чаще встречается двухбугорковый. Увеличение робустности премоляров, отсутствие передних дополнительных бугорков на р2 и р3, редукция талонида и сокращение количества бугорков на нём, а также отсутствие метаконида на m1 у пахикрокут, очевидно, являются прогрессивными признаками. К таковым относятся образцы из Странска скалы, Гомбасцога, Петралоны, Зюссендорна, Конепрусских пещер и др.. В Греции P. brevirostris известна как из поздневиллафранкских местонахождений (Gerakarou 1 – соместно с Pliocrocuta perrieri, Livakkos), так и из эпивиллафранкских (Apollonia 1, Kalamoto, ?-Petralona). Материал из Аполлонии представляет фрагменты с крупными и массивными хищническими зубами – Р4 с относительно сниженным протоконом, высокой коронкой с большими дополнительными бугорками, а также удлинённым и широким m1 без метаконида и коротким однобугорковым талонидом. 
Pachycrocuta brevirostris, Каламото (Kalamoto), Греция: 1 – краниальный фрагмент с нижней челюстью (KLT 201), 2 – правая ветвь нижней челюсти (KLT 215)
Образцы из Каламото демонстрируют сходную морфологию: Р4 с массивным протоконидом, рудиментарным метаконидом и m1 с однобугорковым талонидом. Упоминание этого вида в фауне среднего плейстоцена Петралоны проблематично – описаны лишь фрагменты молочного Р2 и Р4 – чуть крупнее, чем максимальные значения Р. perrieri. С другой стороны, морфология и размер зубов согласуются с Р. brevirostris. 
Нижнечелюстные фрагменты Р. brevirostris (№ 296, 297, 298) из Манастирек (Manastirec), Македония
Гнатодентальные фрагменты известны и из среднеплейстоценовой локации Манастирек (Manastirec), Македонии. Размеры зубов сопоставимы с данными по Р. brevirostris, р2 сравнительно крупный, с высокой коронкой, большим протоконидом и редуцированной задней вершинкой; р3 с небольшим задним бугорком и цингулумом; передние и задние бугорки на р4 являются высокими и не столь чётко отделены от протоконида, как у Crocuta, m1 утолщённый с очень широким тригонидом и небольшим талонидом.
Выводы:
Поздневиллафранкский, раннеэоплейстоценовый интервал (зоны MN 18 - MQ 19) в Европе изучен достаточно хорошо, в целом хорошо отделяясь от позднеплиоценового. Комплекс хищных млекопитающих этого периода наиболее полно представлен в местонахождениях Сенез и Блассак во Франции, Оливола, Верхнее Вальдарно в Италии, Эрпфинген в Германии, Кишланг в Венгрии и др. Одним из самых богатых местонахождений поздневиллафранкского возраста является Оливола в Италии. Интересно, что здесь наряду с архаичной Pachycrocuta perrieri впервые появляется Pachycrocuta brevirostris. Совместные находки этих двух форм обнаружены также в германской локации Эрпфинген и в Верхнем Вальдарно, хотя эти фауны стратиграфически несколько моложе. В Центральной Европе богатый по составу комплекс хищников наряду с P. brevirostris представлен в местонахождении Кишланг, стратиграфически коррелируемый с хапровской фауной Восточной Европы.
Позднеэоплейстоценовый комплекс хищников (зона MQ 20) представлен в местонахождениях Сензель и Валлоне во Франции, Фоггия в Италии и представителями таманской фауны в России. Во всех этих местонахождениях присутствуют ассоциации хищных млекопитающих, также имеющих отличающийся от виллафранкского состав представителей.
Фауна Европы конца раннего – начала среднего плейстоцена (зона MQ 21-22) хорошо изучена. Наиболее интересны для сравнения фаунистические комплексы Западной Европы в местонахождениях Зюссенборн, Странска Скала, Гомбасцог, Петралона. Наиболее характерными элементами этого комплекса хищников в Европе становится крупная P. brevirostris, дентальная морфология которой несколько отличается от поствиллафранкской. В Центральной и Северо-Восточной Азии крупная пахикрокта, близкая к P. brevirostris, известна из местонахождений Тологой на р.Селенга и Засухино (Забайкалье), из илийского териокомплекса Казахстана локации Актогай на р.Чарын, Алма-Атинская обл. и из Лахути 2 на р. Обимазар Таджикистана.
В описании прогрессивной формы Р. robusta Кретцой (Kretzoi, 1938) разделяет европейских пахикрокутовых на две группы: к первой относятся экземпляры, у которых разность между длинами р4/m1 существенно больше. Однако при измерениях в работе Хауэлла и Петтера (Howell & Petter, 1980) явная разница не выявлена, хотя и несколько заметна, но это может являться результатом хроногеографических популяционных вариаций таксона. Также имеется небольшая изменчивость в р4 относительно длины парастиля по отношению к паракону. Более диагностична и заметна изменчивость в морфологии m1: а) число бугорков талонида; б) наличие/ отсутствие метаконида; в) высота параконида и протоконида.
Так, морфотип m1 с двухбугорковым талонидом характеризует, в основном, виллафранкские формы, в то время как однобугорковый, в большинстве, своём, у пост-виллафранкских образцов Р. brevirostris. Куртен и Гаревски (Kurten & Garevski, 1985) отмеча¬ют, что во многих случаях талонид m1 у пахикрокуты имеет единственный режущий бугорок и такое строение является типичным для этой гиены, но у ранних форм встречаются варианты с двумя и тремя бугорками на талониде. В целом исследования показывают, что у Р. brevirostris из фаун галерия (конец раннего - средний плейстоцен) преобладает m1 с одним бугорком? в то время как у поздневиллафранкских форм чаще встречается двухбугорковый талонид. Увеличение робустности премоляров, отсутствие передних дополнительных бугорков на р2 и р3, редукция талонида и сокращение количества бугорков на нём, а также отсутствие метаконида на ml в эволюции рода Pachycrocuta, по-видимому, должны рассматриваться как прогрессивные признаки.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#677 09 September 2016 20:02:50
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Общие выводы:
1. Центр происхождения плио-пахикрокутовых гиен до сих пор неизвестен ввиду отсутствия адекватных ископаемых остатков на границе миоцен-плиоцена после вымирания Adcrocuta eximia. В то время как некоторые исследователи рассматривают гипотезу африканского происхождения, другие полагают, что изначально пахикрокутовые эволюционировали в Азии. Также проблематичен вопрос и о предковых формах – одни полагают изначальными формами африканских Ictitherium (Palhyaena) , тогда как другие считают, что пахикрокуты отделились от некоей ветви азиатских Thalassictis, у которых уже наблюдается тенденция к расширению и большей массивности премоляров. Следует признать, что обе этих гипотезы несостоятельны, так как данные таксоны принадлежат к группе гиеноморфных Ictitheriidae, филогенетически весьма отличающейся от истинных Hyaenidae.
2. В общем плане плио-пахикрокутовые демонстрируют существенные дентальные отличия в эволюционном развитии как от Hyaena, так и от Crocuta - данная апоморфная специализация позволяет позиционировать Pachycrocuta отдельным родовым таксоном вкупе с родом Pliocrocuta. У гиен этой группы длина нижнего хищнического зуба значительно больше длины последнего нижнего премоляра. Подобный признак может рассматриваться у пахикрокут как архаичный, к тому же увеличение ширины и массивности премоляров наблюдается у плио-пахикрокут от плиоценовых P. pyrenaica и Р. perrieri к эоплейстоцен-раннеплейстоценовой P. brevirostris.
Строение m1 является одной из основных характеристик - здесь можно выделить несколько морфотипов строения нижнего хищнического зуба:
I - m1 с выраженным метаконидом и двухбугорковым талонидом;
II - m1 с редуцированным метаконидом и двухбугорковым талонидом;
III - m1 без метаконида с двухбугорковым талонидом;
IV - m1 с редуцированным метаконидом и однобуторковым талонидом;
V - m1 без метаконида с однобугорковым талонидом.
Причём между морфотипами I, II и III, характерными для pyrenaica/perrieri и морфотипами IV и V, характерными для P. brevirostris, в европейских локациях не наблюдается постепенного перехода, что объясняется более поздней инвазией P. brevirostris из азиатских регионов и её эволюцией вне Европы. Однако в целом филетическая связь между раннеплейстоценовой Pliocrocuta pyrenaica, позднеплиоцен-раннеплейстоценовой P. perrieri и эоплейстоцен-среднеплейстоценовой Pachycrocuta brеvirostris заметна в изменениях дентальных пропорций, постепенной утрате метаконида на нижнем хищническом зубе, сокращении талонида и уменьшении количества бугорков на нём. Причём подобная тенденция наблюдается как в европейских популяциях (эпивиллафранкские и среднеплейстоценовые пахикрокуты), так и в азиатских формах - имеет двухбугорковый талонид, тогда как Р. sinensis – однобугорковый.
3. Родословная группы начинается с русцинийской Pliocrocuta pyrenaica – формы, известной из французских местонахождений Перпиньян, Лайна, испанской ла Калера и Одесских катакомб. Однако здесь следует учитывать и азиатские находки из Удунги, свидетельствующие, что ареал пиренейской гиены не ограничивался средиземноморским регионом. Кроме того не исключено, что образец, описанный как Hyaena praecusor из Айн Бримба (Ain Brimba) Туниса, также относится к этому виду (Randall, 1981). В целом дентальная морфология представляет собой более примитивное состояние, чем у Р. perrieri.
4. В предвиллафранский период начала формироваться Pliocrocuta perrieri – дифференциация этого таксона происходила в Азии, а находки в Аль эль-Оуглэм свидетельствуют о довольно широком его распространении. В виллафранке Р. perrieri стала доминантным гиенидом в западноевропейских и циркумсредиземноморских фаунах с ареалом, охватывающим Западную Азию до северного Китая.
Ряд авторов синонимизировали Pliocrocuta с Pachycrocuta, однако морфологический и кладистический анализ говорит о сестринском положении этой формы. Также предлагалась синонимия с современной бурой гиеной, Hyaena brunnea (Turner, 1990). Тем не менее, наличествуют явные признаки, особенно в форме затылочной области, которые чётко отличают эти таксоны в той степени, что синонимия крайне маловероятна.
В конце эоплейстоцена и раннем плейстоцене количество перьерских гиен сокращается и к концу раннего плейстоцена плиокрокуты вымирают – по всей вероятности, в связи с появлением Pachycrocuta brevirostris.
5. Происхождение Pachycrocuta остаётся неясным, так как этот вид появляется почти одновременно 3,0 млн лет назад в Африке и Азии. Однако, несомненно, что пахикрокута эволюционировала в качестве боковой филогенетической ветви неких популяций Р. perrieri где-то за пределами Европы и, наиболее вероятно, в локальных регионах Азии. Подобный крупный хищник должен был сформироваться в условиях открытых ландшафтов с очень высокой экологической продуктивностью в ассоциации с крупноразмерными копытными. Рассредоточение Pachycrocuta в Европе имеет чёткий азиатский контекст, так как она появляется почти одновременно с крупными полорогими (Leptobos, Eucladoceros). Морфологическое сходство между Pliocrocuta и Pachycrocuta позволяет предположить наличие единой эволюционной линии, характеризующейся утолщением премоляров, а таже сокращением талонида и метаконида на m1 (Olive, 2006). В тоже время нижний хищнический зуб у пахикрокуты имеет более выраженный режущий тип, дробящие структуры которого сильно редуцированы - следовательно, у Pachycrocuta по сравнению с Pliocrocuta наблюдается специализация зубного аппарата как для дробления костей, так и для разрезания мяса, что проявляется и в морфологии молочных коренных.
Pachycrocuta изначально была очень успешным таксоном, распространившимся на обширной территории и дифференцировавшимся на ряд подвидов. Подобная дифференциация могла происходить как в периоды миграции, так и локально. В Азии пахикрокута известна из позднеплиоценовых горизонтов местонахождения Нихэвань, где описана форма Р. licenti. Впоследствии она распространяется на юго-восток и наличествует в териокомплексах до среднего плейстоцена – яванская форма P. bathygnatha (Geraads 1979) и P. synensis из локации Чжоукоудянь Китая (Pei, 1934). Африканская Hyaena bellax также отнесена к роду Pachycrocuta, хотя подобная синонимия всё ещё под вопросом. Верделин и Солюниас (Werdelin & Solounias, 1991) считают, морфологические характеристики африканской формы более примитивны, чем у евразийской, в то время как эти различия могут рассматриваться и в качестве индивидуальных вариаций (Turner & Anton, 1996). В любом случае, при отсутствии сравнительного анализа P. brevirostris / P. bellax этот вопрос остаётся открытым. В конце виллафранка пахикрокута появляется и в Европе, где сосуществовала с P. perrieri во время первого континентального оледенения. В кромерское межледниковье (0,7-0,5 млн лет назад) Pachycrocuta вымирает – по-видимому, в Европе чуть раньше, чем в Азии.
6. Таксономический статус средне-позднеплейстоценовой Pliocrocuta (Hyaena) prisca до сих пор проблематичен. Некогда её рассматривали в контексте родственных связей с полосатой гиеной Hyaena hyaena. Также морфологически она схожа с бурой, H. brunnea, но несколько крупнее, и отличается от этого вида рядом диагностических признаков. Подобная форма известна из Палестины во время эемского межледниковья и вюрма. Предполагается, что новое появление Pliocrоcuta в Европе в среднем плейстоцене представляет собой ререколонизацию из Азии, так как её отсутствие в других местонахождениях на Пиренейском полуострове после исчезновения в виллафранке противоречит миграции из Португалии. Кроме того, не исключено, что на самом деле эта форма является поздним реликтом рефугиальных популяций Pliocrocuta, впоследствии диспергировавших в другие регионы.
7. Pachycrocuta была самым крупным когда-либо существовавшим гиенидом, хорошо приспособленным для утилизации мяса и потребления костей. Помимо своего огромного размера, Р. brevirostris отличалась от современных гиен укорочением дистальных сегментов конечностей, в результате чего их общая высота была не намного больше, чем у крупных экземпляров современной пятнистой гиены. Кроме того, это указывает на меньшие беговые качества хищника. Тем не менее, имеется мнение (Dennell et al., 2008), что огромный размер P. brevirostris мог быть быть выгоден при охоте на крупноразмерных копытных или в случае конкурентности с другими хищниками. В любом случае, стоит отметить, что поведение ныне живущих хищников не могут быть в полной мере экстраполированы непосредственно на ископаемые таксоны.
Оценки размеров P. brevirostris получены с использованием уравнения регрессии, скорректированных с современными хищниками. В частности, наиболее точная оценка обеспечивается уравнением Андерссона (Andersson, 2004), в котором масса вычисляется по периметру дистального отдела плечевой кости. Во всяком случае, уравнение даёт оценки размера для современных гиен, очень близкие к их фактической массы. Для Р. brevirostris оценки получаются следующие: по экземпляру плечевой кости из Инкарнал (116,3 мм) вес животного составлял 116,6 кг, по плечевой кости из Дманиси (104,4 мм) - 104,4 кг, что обеспечивает среднюю массу 110,5 кг. Пропорции конечностей предполагают высоту в плечах до 1 м (Turner & Anton, 1996).
Биомеханика черепа P. brevirostris демонстрирует, что гигантские гиены имели значительные механические преимущества для костедробления благодаря массивности премолрной серии, хорошо развитым челюстным мышцам (особенно жевательным и крыловидной) и глубоким нижним челюстям, хорошо приспособленных для высоких нагрузок. В общем плане, реконструктивно P. brevirostris выглядит как крупный хищник, достигающий размеров львицы, с массивными конечностями с укороченными дистальными отделами и тяжёлыми, массивными челюстями с хорошо развитым зубным аппаратом. Все эти особенности отражают адаптацию к утилизации трупов копытных, транспортировке больших кусков и костедроблению для извлечения костного мозга.
8. Первый этап видообразования плио-пахикрокутовых гиен был обусловлен похолоданием климата и исчезновением крупной Adcrocuta. Второй этап соответствует большему охлаждению в конце русциния и начале виллафранка, что стимулировало распространение Pliocrocuta. Третья, раннеплейстоценовая, фаза датируется 1,7 млн лет назад, с широким распространением рода Pachycrocuta. Этот период конца виллафранка отличается от двух предыдущих появлением крупноразмерных полорогих: Bos, Bison, Ovibos и Megaceros и исчезновением более половины таксонов. Исчезновение гигантской короткомордой гиены также является частью комплексного вымирания 0,5-0,4 млн лет назад наряду с Рanthera gombaszoegensis.
Стратиграфическое распределение Р. brevirostris предполагает, что это произошло в первом или втором этапе миндельского оледенения. Её последние находки фиксируются в местонахождениях начала среднего плейстоцена – Зюссенборне, Фоггии, на Британских островах и др. Однако форма, сходная с перьерской гиеной, вновь появляется в среднеплейстоценовых локациях – Мауэр, Мосбах, Эскель и Люнель-Виэль, где она идентифицируется как Pliocrocuta (Hyaena) prisca. Интересно, что эта дисперсия Pliocrocuta выглядит несколько ограниченной, так как она не регистрируется в Италии или Великобритании.
Видообразование во время максимальной ледниковой фазы среднего плейстоцена привело к эволюции специализированной Crocuta spelaea и конкуренция с пещерной гиеной вполне может быть одним из факторов, которые способствовали исчезновению пахикрокуты. Оба вида сосуществовали в течение короткого периода и сравнение показывает, что Pachycrocuta brevirostris конкурировала с Crocuta praespelaea, которая имела более широкий и пропорционально крупный р3, обеспечивающий более эффективное костедробление. К тому же размеры и пропорции хищнических зубов также указывают на более развитую функцию разрезания мяса у пещерной гиены. Поэтому представляется оправданным, что Crocuta кажется, удалось потеснить Р. brevirostris на всём ареале. Возможно, немаловажную роль в этом сыграть и мосбахский лев, Panthera fossilis, в чей рацион входили копытные того же габаритного диапазона.
В то же время не совсем ясно сосуществование Crocuta и Pliocrocuta (Hyaena) prisca, отмеченное в Люнель-Виэль и ряде местонахождений на Пиренейском полуострове. Не исключено, что второй вид, будучи меньшиих размеров, занимал другую экотрофическую нишу, не представляя особой конкуренции.
Окончательные причины вымирания всегда сложны и многозначны. Адаптивные различия между Pachycrocuta и Crocuta хорошо отражены в морфологии и типе износа зубных рядов. С другой стороны, вымирание пахикрокут могло произойти не в результате конкуренции и главным фактором было изменение окружающей среды и климатические колебания.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#678 09 September 2016 20:07:08
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Вроде бы, бОльшую и наиболее интересную часть фоссильных находок плио-пахикрокут рассмотрел… Не всегда, правда, фотками иллюстрировал, звиняйте. Кому интересно – обращайтесь. Чем могу, всегда рад поделиться.
Как всегда, приватные выводы:
1. Исследователями подозревается о явной связи азиатских и африканских пахикрокутовых. Однако почему-то никто до сих пор не сделал фундаментальных выводов о том, насколько африканские плиоценовые пахикрокуты морфологически идентичны ранним формам евразийских плиокрокут. И почему китайских пахикрокут плиоцена буржуи рассматривают именно как Pachycrocuta, а вот наши учёные гиену из Удунги видят как Pliocrocuta, которая конспецифична ранним формам плио-пахикрокутовых? Причём справедливо отмечая, что среднеплейстоценовая гиена из Тологоя – уже образчик прогрессивной пахикрокуты, а не плиокрокута.
Также игнорируется мнение русских учёных по среднеазиатским формам. Например, в Лахути - Pliocrocuta, а в Куруксае уже Pachycrocuta, хотя разница в возрасте не очень большая.
2. Отсутствует общее мнение о связи Crocuta sivalensis с сиваликской популяцией Pachycrocuta (описываемой как Р. brevirostris neglecta, однако геохронологически и по некоторым морфопризнакам она всё же ближе к группе sinensis) – оба таксона практически сосуществовали и некоторые таксоны (C. colvini, C. felina) некоторые палеонтологи синонимизировали то с Crocuta, то с [i]Pachycrocuta, но однозначного мнения так и нет. Ревизия по гиенам Восточной Азии просто плачет горючими слезами.
3. Также весьма проблематична атрибуция позднеплиоценовых китайских крокутиин. Данные гиены – аппендиксная популяционная ветвь перьерских плиокрокут или примитивная ветвь пахикрокутовых, таксономически обозначенная как Pachycrocuta licenti? Или всё же там присутствовала эволюционная дифференциация? Судя по образцам, это вроде наблюдается, но ведь серии нет.
Серия китайских пахикрокут, причём изумительная (около двухсот номерных образцов черепов, нижних челюстей и даже несколько полных посткраниальных скелетов) есть только в Нихэвань, но там уже явная позднеплиоценовая Pachycrocuta sinensis.
4. Пресловутая Hyaena (=Pliocrocuta) prisca на границе среднего-позднего плейстоцена - вот чует сердэчко, «винегретный» таксон. «Раздербанится» он со временем на представителей реликтовых популяций Pliocrocuta, регрессивных эндемиков из Pachycrocuta, а некоторые образцы вообще припишут ранним формам Crocuta spelaea.
5. По поводу разделения рода Pachycrocuta на виды. Я сам ничего не придумывал, а только руководствовался хронологией находок, сравнением фаунокомплексов, сравнительной морфологией в описаниях, географической зональностью и обыкновенной логикой:
- Pachycrocuta bellax, «слепая» филогенетическая «веточка» из эволюционного ядра азиатских плиокрокут, мигрировавших на запад – очевидно, диффузировавшихся популяционно в Малой Азии, но в Африке эндемично приобрётших специфические характеристики;
- Pachycrocuta licenti, пахикрокута северо-восточной Азии (Монголия и Китай). Не исключено, что мозаичный таксон, фактически представляющий как прогрессивных перьерских плиокрокут (м. Шамар), так и примитивных пахикрокутовых (м. Нихэвань), или же региональная переходная форма, но основе которой сформировался более продвинутый таксон – последняя версия поддерживается русскими палеонтологами;
- Pachycrocuta sinensis - по сути, также проблематичная «разновидность», так как её часто синонимизируют с типовым видом brevirostris. Однако имеется ряд отличий не только помимо морфологических, а и на основе логики эволюционного генеза. Имеется подвид bathygnatha в среднем плейстоцене Явы, и это пахикрокута. Однако эта форма гнато- и дентоморфологически гораздо архаичней европейской и ближе к группе sinensis. Отсюда вывод, что sinensis – «узловой» таксон, а значит brevirostris, хоть и сателлитный вид, но не настолько близкородственный;
- Pachycrocuta brevirostris, вот здесь без особых проблем, типовая европейская форма одного из доминантных хищников ранне- и среднеплейстоценовых биомов Европы. Ну, здесь, набравшись наглости, могу заметить, что данную форму для удобства можно разделить на два хроноподвида: ранне- и среднеплейстоценовый по морфологии талонида, на это и Марина Владимировна внимание обращала (Сотникова и др., 2002).
6. В линии плио-пахикрокутовых гиен наличествовало несколько миграционных «волн». Возможно, реверсивных:
А). Первая, русцинийская по датировке фаунокомплексов Европы (примерно 5 млн лет назад), выделенная там как Pliocrocuta pyrenaica (Serrat d’en Vacquer, Layna, Одесские катакомбы), также присутствующая в Азии (Удунга) и, не исключено, в северо-восточной Африке (Kanapoi, Ain Brimba).
Б). Вторая форма - Pliocrocuta perrieri, которая экоморфно начала уверенно занимать нишу костедробящих падалеядов после вымирания Adcrocuta – возможно, ввиду постепенной смены териофауны или более продвинутой социальной организации. Перьерские плиокрокуты уже более широко распространены – по всей Азии и Европе вплоть до Великобритании и уверенно занимают экологически благоприятный средне-переднеазиатский «пояс». Также в среднем плиоцене проникают в северо-восточные регионы Африки.
В). В позднем плиоцене Азии формируется прогрессивная пахикрокутовая «морфа». Прохорезно «выстрелившая» (да простят меня академики) на запад 3,5 млн лет назад и аппендиксно «завернувшая» в Африку, проникнув аж до юга, а в Азии – в Индонезию…
Но здесь – бифуркационная точка Pachycrocuta и собственно Crocuta. Если эволюционный центр последней (плиоценовые Crocuta eturono, C. dietrichi) вроде бы общепризнан в Африке, то пахикрокутовые под вопросом – но, очевидно, всё же в Азии. И сходство обусловлено скорее конвергенцией.
Г). Pachycrocuta brevirostris, сменяющая в Европе Pliocrocuta и, в свою очередь, в позднем плейстоцене сдающая позиции Crocuta.
И здесь не только анализа ранних крокутоидных гиен с пахикрокутными формами никто не проводил, а и никто не сравнивал ранних (позднеплиоценовых) пахикрокут Африки и Азии. Как и масштабно не сопоставляли архаичных пахикрокут с прогрессивными плиокрокутами. Единственный толковый, достаточно объёмный и много что аргументированно объясняющий вывод имеется у МВ Сотниковой на анализе азиатского материала. Африканский упоминается, но не анализируется, но это этически и профессионально понятно – если непосредственно с образцами не работаешь, зачем быть голословным?
Примерно 1,8-1,6 млн лет назад Pachycrocuta радиировала в Европу (номинативная форма), где успешно существовала до 0,5 млн. Согласно исследованиям (Ficcarelli & Тоrrе, 1970 и др.), европейская пахикрокута не демонстрирует явных изменений, хотя выглядит странным, что на протяжении миллиона лет таковые не проявляются. Однако некоторые признаки наличествуют – морфология талонида (у большинства среднеплейстоценовых образцов однобугорковый и двухбугорковый у раннеплейстоценовых), хотя данная характеристика вариативна; а также морфометрические данные: пропорциональное соотношение длина/ширина р4 и m1, а также длина р4/ m1 у среднеплейстоценовых, укладываясь в размерный диапазон раннеплейстоценовых, скорее группируются в нижних значениях последних. Всё это. в принципе, позволяет предположить о наличии двух форм пахикрокут – эпивиллафранкской и среднеплейстоценовой хотя бы на уровне хроноподвидов.
Обратившись к палеонтологическим свидетельствам, примем как должное, что пахикрокуты на протяжении среднего плейстоцена занимали одни из доминантных трофических позиций в экосистемах Палеарктики. В итоге в Африке они были вытеснены современными пятнистыми гиенами в конце раннего плейстоцена, а к концу среднего плейстоцена в евразийском регионе – спелеоидными.
Pliocrocuta (Hyaena) prisca всё же, очевидно, является реликтовой формой. Логичней считать её потомком архаичных плиокрокут, нежели собственно гиен или пахикрокут. А редуцированность метаконида на m1, являющаяся признаком прогрессивности – вариативная характеристика, нередко проявлявшаяся и у перьерских гиен, так что обособленные рефугиальные популяции, распространившиеся после исчезновения пахикрокуты, вполне могли его сохранить. Жаль, что этому морфопризнаку не уделено должное внимание в статьях по описанию серии образцов Р. prisca из Португалии (Furninha).
7. Мне нравится разделение на трибы, оно более глубже отображает филогенетические связи в классической систематике. И в ней плио-пахикрокутовые должны быть выделены в отдельную группу, занимающие сестринское положение к собственно крокутинам.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#679 09 September 2016 20:08:21
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Возможно, при чтении текста с п. 668 в глазах рябит не столько от дентологических терминов (хотя я старался диагностику максимально упростить и не все сравнения или соображения по «зубкам» выложить, это мало кому интересно (Глеб Анатольевич, старик, помню твои замечания – кстати, матушке, как всегда, низкий поклон, какая изумительная Женщина!)), сколько от обилия «КРОКУТА». Ну, тема такая, братцы, извините… Для иллюстрации и вящего осознания всего вышеописанного представляю филогенетическое древо плио-пахикрокут, выросшее на данный момент в моём понимании: 
Ну, и напоследок, личное и несколько «оффтопное» мнение. Инициированное не оголтелым патриотизмом, а беспристрастной объективностью. А посему считаю, что «ядро» здравомыслящего и наиболее компетентного взгляда на семейство Hyaenidae в настоящий момент сосредоточено в Восточной Европе в лице Семёнова, Барышникова и Сотниковой. В Западной единственный адекватный чел – Дидрих (был ещё Бьорн Куртен, но мало прожил, всего 64 года, многого не успел). Работы всех остальных интересны лишь с точки зрения описательной и сравнительной части. Глубоко уважаю Верделина, однако после монографии «The Hyaenidae taxonomy, systematics and evolution» к мнению дядьки Ларса относительно гиен, наверное, всё же стоит отнестись скептически. И после его коллективной статьи с Антоном и Тёрнером «Taxonomy and evolutionary patterns in the fossil Hyaenidae of Europe», в которой они, к примеру, предполагают синонимию пахикрокутовых с ликиеной (?!), да и вообще рассматривают плиокрокут как крупную форму ископаемой гиены, близкой к современной полосатой (что заметно по антоновским иллюстрациям), градус предвзятости только повысился.
Да, ещё - у буржуинов были ещё Хауэлл и Петтер. Старенькие работы, но сколько в них новаторских мыслей и интуитивного предвидения - вот это, безоговорочно, «молотки»! Дуэт покруче, чем «Modern Talking» ![]()
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#680 18 February 2017 13:35:01
- Алекс
- Сан Саныч - Модератор

- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические гиены
 Чей синоним гиена арверненсис ? череп 35 см ,как у крупного льва !
Чей синоним гиена арверненсис ? череп 35 см ,как у крупного льва !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#681 19 February 2017 14:59:12
- shish02
- Гость
Re: Доисторические гиены
Алекс :
http://savepic.ru/12948258m.png Чей синоним гиена арверненсис ? череп 35 см ,как у крупного льва !
Если не ошибаюсь, сейчас это Pliocrocuta perrieri. Во всяком случае, в работе Барышникова и Верещагина "Краткий обзор четвертичных гиен (семейство Hyaenidae) России и сопредельных территорий гиена арверненсис указана как синоним Pliocrocuta perrieri
#682 19 February 2017 15:27:38
- shish02
- Гость
Re: Доисторические гиены
Да и у Верделина и Солоуниаса синоним H. arvernensis - Pliocrocuta perrieri
#683 19 February 2017 15:36:29
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические гиены
shish02 :
Да и у Верделина и Солоуниаса синоним H. arvernensis - Pliocrocuta perrieri
Спасибо . Выходит она в одном размерном классе с пахикрокутой !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#684 21 February 2017 23:11:56
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Овернская гиена - синоним перрьерской формы плиокрокуты.
Алекс :
Выходит она в одном размерном классе с пахикрокутой !
В принципе да, хотя пахикрокута, как представляется, плиокрокуты примерно процентов на 20 крупнее была.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#685 22 February 2017 00:56:57
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические гиены
Сorvin :
Овернская гиена - синоним перрьерской формы плиокрокуты.
Алекс :
Выходит она в одном размерном классе с пахикрокутой !
В принципе да, хотя пахикрокута, как представляется, плиокрокуты примерно процентов на 20 крупнее была.
 По наибольшим черепам пахикрокуты -36 см общая длина ,и плиокрокуты -35 см, разницы особо нет ,разве что плиокрокута имела относительно тела крупную голову.
По наибольшим черепам пахикрокуты -36 см общая длина ,и плиокрокуты -35 см, разницы особо нет ,разве что плиокрокута имела относительно тела крупную голову.
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#686 22 February 2017 10:01:49
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Саныч, это, безусловно, аргумент, но попробуй ещё сравнить у этих же образцов соотношение (кондилобазальной !) длины с шириной (скуловой или между карнассиалами). А ещё лучше, для чистоты эксперимента - у нескольких экземпляров.
Да и зубки у пахикрокуты всё же крупнее были.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#687 22 February 2017 11:06:15
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические гиены
Сorvin :
Саныч, это, безусловно, аргумент, но попробуй ещё сравнить у этих же образцов соотношение (кондилобазальной !) длины с шириной (скуловой или между карнассиалами). А ещё лучше, для чистоты эксперимента - у нескольких экземпляров.
Да и зубки у пахикрокуты всё же крупнее были.
По КБ да ,разница в 4 см ,а вот по ширине черепа относительно КБ у плиокрокуты шире . Выборка черепов пахикрокуты больше ,поэтому среднее значение будет не совсем достоверным .
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#688 22 February 2017 19:11:22
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Не, Саш, серия как раз и нужна. Я тебе лучше в личке отпишусь.
Я как-то по этой группе приватное исследование проводил – правда, не в должной степени серьёзное, как хотелось бы, но пару месяцев «повтыкал». Морфологически они довольно схожи и, по сути, составляют одну кладу. Однако представляется, что Pachycrocuta всё же была эволюционно прогрессивней и экологически успешней, иначе она бы плиокрокутку не вытеснила. Так же, как тихой сапой Pliocrocuta «съела» адкрокуту, а ведь Adcrocuta тоже солидной зверюгой была…
Саныч, да иногда тупиковые вопросы возникают не только по глобальным проблемам типа вымирания динозавров, плейстоценовой мегафауны или саблезубых котов, а и по таким - вот почему пахикрокуты вытеснили плиокрокут? Я даже аналогии навскидку привести не могу… Львы появляются в голоценовой Европе после явной сдачи границ ареалов пещерными пантерами, плейстоценовые леопарды выживают виретайлуров ввиду более явной экопластичности и т. д. – тут более-менее ещё к чему-то «привязаться» можно. Даже по рысям – иссуэрские рысеподобные кошки сужают ареалы распространения после серии среднеплейстоценовых (гюнц-миндель, миндель-рисс) оледенений, и их место занимают более адаптированные линксоподобные фелины.
Пока представляется постепенная замена более эволюционно продвинутыми формами – сформировавшимися в том же региональном ядре и радиировавшими теми же миграционными путями. Но это лишь теория.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#689 20 April 2017 22:35:22
- Crazy Zoologist
- Гость
Re: Доисторические гиены
Адкрокута.
#690 21 April 2017 22:05:47
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Артём, спасибо! Клёвое фото. И личная просьба - на подобные фото можно ссылки?
А теперь ложка дёгтя - адкрокута весьма проблематичный таксон. Её до сих пор аргументированно не могут никуда "внедрить". Дело в том, что морфологически это гиеноподобный хищник, но современные гиены появляются около 6 млн лет назад; в позднем туролии – группа гиенотериев (пусть и хвастаюсь, уж простите, но ряд нижних зубов с клыком до моляра гиенотерия собственноручно с Серёгой Куршаковым извлекал), которые филогенетически вроде бы и являются предками современных гиен.
Но, позвольте, гиенотерии – 6 млн и это довольно примитивные и морфологически генерализованные хищники. А диапазон существования адкрокуты : 10-6 млн, т. е. гораздо раньше.
Однако тогда существовали и другие гиенофелиморфные хищники типа перкрокутовых или иктитериев.
А принадлежит ли адкрокута к истинным «гиенам»?
P.S. АртёмСаныч, дай ссыль на фото, такое упускать нельзя. Заранее спасибо.
И, старина, топливо в горнило твоих идей - попробуй хронологически проанализировать время и географию распространения перкротут, иктитериев, гиен и, отдельно - адкрокут.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#691 21 April 2017 22:30:09
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические гиены
 мощные челюсти у адкрокуты ! Игорь , ссылок на скелет полно ,в основном на китайские ресурсы и википедию !
мощные челюсти у адкрокуты ! Игорь , ссылок на скелет полно ,в основном на китайские ресурсы и википедию !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#692 21 April 2017 22:50:59
- Crazy Zoologist
- Гость
Re: Доисторические гиены
Игорь, фото взято из всеми известной википедии:-).
Там же указано (с приведением источника - Macdonald 1992), что гиены разделились на костедробящих и собакоподобных примерно 10-12 млн лет назад. Так что адкрокута вписывается в эти хронологические рамки.
#693 21 April 2017 22:51:54
- Crazy Zoologist
- Гость
Re: Доисторические гиены
P.S. И определите наконец-то костяху, которую я выложил в псовых!:-)
#694 21 April 2017 22:57:29
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Для меня Вика не авторитет и в неё редко заглядываю.
ОК, братцы, укол принят... А вот "туше" это или нет - дайте время.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#695 21 April 2017 23:37:12
- Crazy Zoologist
- Гость
Re: Доисторические гиены
Я бы сказал, что для меня это не совсем авторитет. Но когда приведены источники, тут я верю. Надо сказать что английской я доверяю больше. Да и потом что такое вика? Перепечатывание статей из разных ресурсов. И могу сказать что ляпов там не больше, чем в науч. популярной литературе.
В данном случае все логично. Около 10 млн лет назад гиенки стали развиваться по двум направлениям. Адкрокута, которая вляется явным костедробильщиком вписывается в эти рамки. Нестычковок не вижу. Ты видишь?
Теперь отсебятина: даже костедробящие гиены отличные охотники как мы видим на примере пятнистых гиен. Хоть и адаптации совсем не охзотничьи:
1. Относительно низкие и очень толстые клыки, при этом тупые.
2. Огромные заклыковые зубы.
3. Малая скорость бега.
Насчет 3-го пунка можно спорить что гиены очень быстры на самом деле, как и гну про которыхз пишут что бегут они со скоростью 80 км в час. Но я очень скептически отношусь к таким заявам, даже в научной литературе. Ну никак не может гну бежать на целых 10 км в час быстрее чем скакун и не может бочкообразная гиена бежать быстрее уиппета. Мой уиппет не самый быстрый, не беговой, но его максимум это 55 км в час, правда по асфальту. Выходит чуть более 55 км в час в рывке. Гиена явно медленнее.
#696 21 April 2017 23:49:37
- Crazy Zoologist
- Гость
Re: Доисторические гиены
Сorvin :
Для меня Вика не авторитет и в неё редко заглядываю.
ОК, братцы, укол принят... А вот "туше" это или нет - дайте время.
#697 22 April 2017 00:15:53
- Crazy Zoologist
- Гость
Re: Доисторические гиены
Гиены ближе к виверрам или мангустам?
http://www.wearesites.com/Personal/Hyen … lution.php
#698 22 April 2017 00:31:53
- Crazy Zoologist
- Гость
Re: Доисторические гиены
Пещерная гиена.

Молодая пятнистая гиена.
Пещерная гиена.
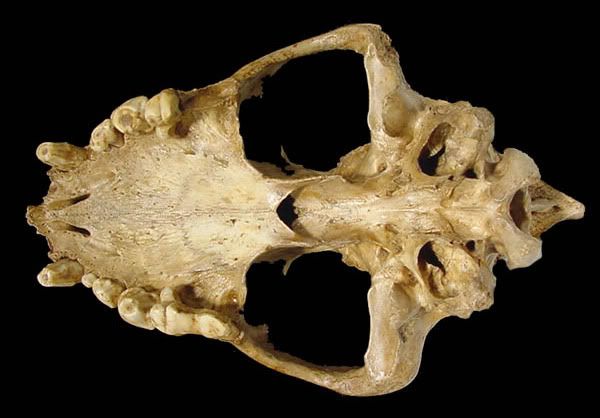
Пятнистая гиена.
Волк.
Лев.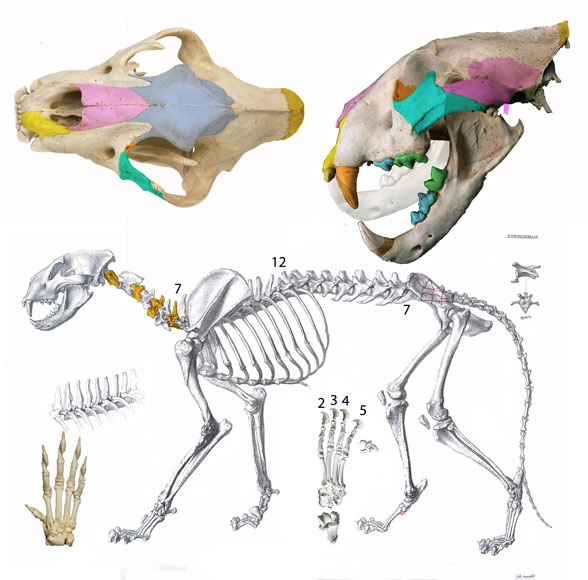
Земляной волк.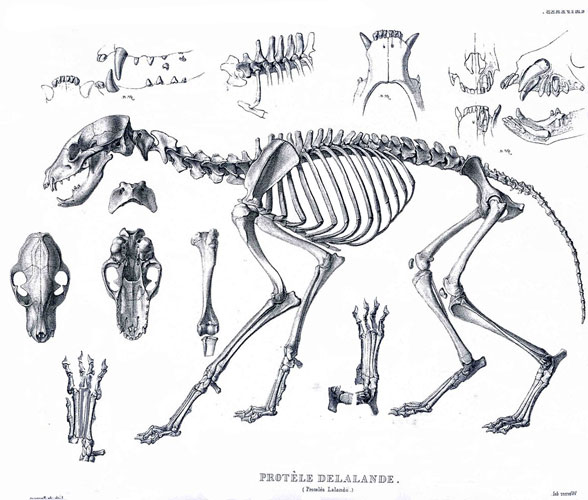
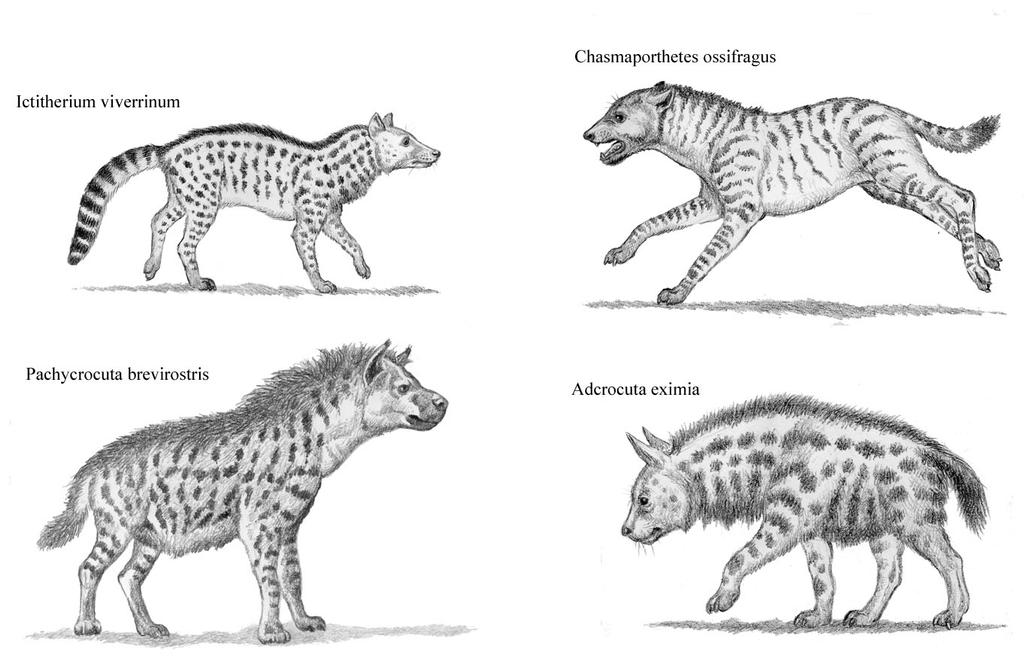
НАПОМНИТЕ МНЕ ПОЖАЛУЙСТА МАССУ ТЕЛА ХАЗМАПОРТЕТЕСА (МИН. - МАКС.).
#699 24 April 2017 01:12:45
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Доисторические гиены
Коллеги, «шпильку» в свой адрес не забыл. Может, иногда я и гипотезирую вразрез с ортодоксальными понятиями, но это вовсе не фантазии.
Что расскажем о семействе гиеновых? Классически – два подсемейства, Protelinae с «земляным волком» и Hyaeninae с крокутами (пятнистая гиена) и гиенами (полосатая и бурая).
Однако на самом деле перед нами – лишь «осколок» обширнейшей группы, экоморфно объединённой адаптивной специализацией.
Братцы, в миоцене появилась новая карниворная группировка среднеразмерных актиных хищников, нашёдшая свою эконишу в плане некоторой приверженности к трупофагии. Причём экологически по этому вектору двигалось много различных линий. Перкрокутиды; чуть позже – иктитерииды и гиениды. Объективно - подобный тренд был присущ не только фелиморфам, вспомните подобное у арктоидов и канид.
Ребят, обожаю хищных млеков, в приватных интересах – фелоиды. Но разрешите Вас уведомить, братцы – гиены не менее интересны, чем кошки. Просто последние внешне более ярки и харизматичны.
И позвольте чуть «поговнячить», рассматривая современные формы…
1. Proteles cristatus, земляной волк – это не гиена, а реликтовая форма перкрокутид. Читаем Chen, Schmidt-Kittler. 1983. «The deciduous dentition of Percrocuta KRETZOI and the diphyletic origin of the hyaenas (Carnivora, Mammalia)» и Барышников, Аверьянов. 1995. «Молочные зубы хищных млекопитающих (отряд Carnivora) - семейства Protelidae и Hyaenidae».
Онтогенез, уважаемые коллеги, тут никуда не денешься… Величайший аргумент – круче, чем базикраниумная топология…
2. Crocuta, пятнистая гиена – все гиениды, демонстрирующие нынешний «крокутоидный» морфотип появляются лишь в конце плиоцена. Все более ранние сходные формы – адаптационная конвергенция, потому что никаких близкородственных филогенетических линий найти невозможно даже при очень большом желании. Ну уж очень большой «разлёт»…
3. Hyaena, бурая и полосатая гиены – более «саркофильный» морфотип. Однако при изучении предыстории этих таксонов явно видно, что это разные роды. Как и Crocuta - гиениды, безусловно. Но проанализируйте их палеонтологическую летопись.
Артём, на хера тебе кишки, старина? Вот будет клёво и действительно полезно, если потратишь время на сравнение репродукционных органов у современных гиенид. О тождестве мужской и женской половой системы у крокут все наслышаны и насмотрены. Но присутствует ли подобное у других гиенид?
P. S. А по поводу адкрокуты палец на курке держу, но нужно хотя бы пару буржуйских букварей «поднять» и освежить в памяти, просто вхолостую брехать не хочется. Помнится, что это палеарктический таксон с евразийскими формами и упоминанием африканской фиксации, морфологически редуцированным протоконом на верхнем карнассиале и т. д. ![]() По-моему, они даже несколько с Chasmaporthetes сходны и, возможно, «предкородственны» с Talassictis. Но талассиктис» насколько разбираюсь в гиенах и как кааца, очень «винегретный» таксон…
По-моему, они даже несколько с Chasmaporthetes сходны и, возможно, «предкородственны» с Talassictis. Но талассиктис» насколько разбираюсь в гиенах и как кааца, очень «винегретный» таксон…
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#700 24 April 2017 12:07:58
- Crazy Zoologist
- Гость
Re: Доисторические гиены
Что-то ты меня запутал. А как же полностью окостеневшая булла? И почему большинство ученых относит его все же к гиенам?