Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#476 22 August 2017 19:42:03
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5935
Re: Эволюция кошкообразных (Feliformia)
А почему бы самому не проштудировать дюжину статей по экологии современных популяций ягуара? И, вооружившись инфой, задать аргументированный вопрос?
Там или кратковременная дискуссия наметится или вопрос сам собой отпадёт...
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#477 22 August 2017 22:34:37
- shish02
- Гость
Re: Эволюция кошкообразных (Feliformia)
Амфицион :
По идее я должен был задать этот вопрос в разделе хищных, но туда никто и знатоков не заходит. А зря. Поэтому спрашиваю тут. Почему ягуар не эффективен в горах?
Не эффективен по сравнению с кем? Если с пумой, то посмотрите на отношение длины передних и задних конечностей этих кошек и все поймете (у ягуаров относительно длинные передние конечности - 89,1 % длины задних, а пумы этот показатель - 81,4 %). Кроме того, ягуар воду любит.
#478 25 August 2017 12:47:36
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Игорь, современный ягуар не показатель, ареал сильно уменьшен, даже со времен голоцена.
shish02, тигр тоже любит и что? К тому же ягуар забирается все же довольно высоко в горы.
#479 25 August 2017 20:44:03
- shish02
- Гость
Re: Эволюция кошкообразных (Feliformia)
Амфицион :
Игорь, современный ягуар не показатель, ареал сильно уменьшен, даже со времен голоцена.
shish02, тигр тоже любит и что? К тому же ягуар забирается все же довольно высоко в горы.
А то, что и тигров в высокогорье нет. Нет воды и леса нет тигров и ягуаров. Все. Под "горами" я в данном случае имел ввиду не только высоту над уровнем моря, но и определенный ландшафт, где густая растительность сходит на нет. В горных лесах (до 2 км в Перу и Венесуэле и даже до 2,7 км в Боливии) ягуар еще обитать может, хотя выше 1 км поднимается редко. Точно так же ведут себя и тигры в Непале (обычно они не поднимаются выше 1,5-1,6 км). Теперь сравните со снежным барсом (в Гималаях летом от 2,7 до 6 км) или с пумой (до 4 км). Кроме того, в своем предыдущем посте вы спрашивали не о том как высоко в горы может забираться ягуар, а о том "почему ягуар не эффективен в горах?" По-моему, двух названных мною факторов (морфологических и экологических), обуславливающих меньшую эффективность в горах ягуара по сравнению с пумой, вполне достаточно для ответа. Хотите большего - последуйте совету Игоря и прочтите сами хотя бы дюжину работ по ягуару.
П/с. Подобного рода вопросы вы уже задавали и даже не один раз. Если это просто способ "оживить" ветку, то это еще куда ни шло. Хотя было бы лучше оживлять ее чем-то новым. Но если вы просто не помните или не воспринимаете ответы на свои вопросы, то все гораздо хуже.
#480 27 August 2017 16:50:26
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5935
Re: Эволюция кошкообразных (Feliformia)
Вот всё же просто донельзя занимает вопрос о гипотетическом эволюционном «ядрышке» фелидоморфных.
- Nimravidae и Nandiniidae: тут истоки, очевидно даже не палеогеном, а палеоценом «пахнут», да и филогенетическая ветка совершенно другая. Единственное замечание: нимравиды в фаунокомплексах эоценовой Азии и Северной Америки появляются почти одновременно, а в Европе датируются МР 21, ранним олигоценом (вполне закономерно, тогда из Европы и самый ранний стеноплезикт известен и наблюдается некоторый фаунообмен с Азией – фиксация тех же антракотериид). Очевидно предположение о наличии некоего азиатского генерационного центра, однако хроногеографическое распространение нимравидин заслуживает более подробного и отдельного поста. Ещё один «вброс» нечто коричневого в вентилятор размышлений: а не могут ли нандинии являться реликтовыми дивергентами нимравид? Безусловно, тут нужен подробнейший анализ;
- Viverridae (как и Prionodontidae– Азия, тут внесомненно;
- Herpestidae (и последовавшие Eupleridae), как вариативные ответвления архаичных виверровых, были распространены в евро-африканском регионе и рекреации, охватывавшей южную половину Азии. Конечно, некоторые филогенетические «побеги» были автохтонны, но ведь начало положили азиаты;
- Ictitheriidae ближе вовсе не к гиеновым, а к герпестидам и эволюционно дистанцировались от древних виверр, очевидно, вместе с последними. Самые ранние остатки – нижний миоцен Причерноморья. Затем находки из миоценовых отложений Западной Европы (Thalassictis, Plioviverrops), позднего миоцена Китая (Sinictitherium) и Африки, а также раннего плиоцена Грузии, Малой и Средней Азии. В отношении иктитериид вырисовывается эволюционная аккумуляция в сарматском регионе (Тетис и Паратетис) на западе средней Азии с последующей популяционной радиацией в восточную Азию и северную Африку, но это первоначальная и очень консервативная гипотеза;
- Hyaenidae на самом деле очень сложное семейство с огромнейшим «белым» пятном. Дело в том, что в миоцене появляется конспецифичная евразийская группа в составе Hyaenotherium, Hyaenictitherium и Miohyaenotherium, вполне претендующая на пьедестал предковой категории для последующих гиенид. Однако таковые в ватаге хазмапортетных и всяческих родов костедробящего морфотипа вовсе не демонстрируют стройность морфологического «ансамбля», т. к. прямой филогенетической связи с данными предками не наблюдается. Некоторое сходство имеется, но оно вполне может быть обусловлено конвергенционным подобием ввиду идентичной экологии. Подробнее об этом можно почитать в монографии Семёнова (1989) по иктитериям, стр. 89-90. А, возвращаясь к «нашим баранам» можно отметить некую азиатскую эволюционную базу, периодически популяционно «выстреливавшей» гиенидами в Европу, Африку и в Пендосию. Естественно, с дальнейшей формацией эндемичных форм, которые позже могли миграционно реверсировать в поисках лучшей доли, но изначальная эволюционная сердцевина от этого не меняется (как и вывеска на борделе после перестановки коек);
- Stenoplesictidae: вот здесь в наличии пока ещё неизвестного очага эволюционной генерации просто уверен. Фиксация морфологически и хронологически близких ранних форм в Европе и Монголии раннего олигоцена (Anictis и Alagtsavbaatar); затем - стратиграфический перерыв в Европе, появление прогрессивных форм в среднем олигоцене Центральной Азии (Shandgolictis) и, тут же - очень схожий с ними типовой европейский Stenoplesictis. Затем – исчезновение стеноплезиктовых в позднеолигоценовой Азии и продолжающееся адаптационное процветание в Европе и Африке. Причём на обоих материках в этой группе наблюдается интересный тренд: прослеживается последовательная филогения в эволюции некоторых эндемичных линий, а вот откуда они возникают, пока неясно. Т. е. примитивные формы (типа Anictis и Viretictis в Европе или Africanictis в Африке) неожиданно появляются в палеонтологической летописи как чёртик из коробки, вне явной связи с сосуществующими таксонами этого же семейства. Ну, типа, если бы сейчас в Ростовской области наряду с кабанами по речным затонам пекари бы вдруг объявились (исключая интродукционное вмешательство). Здесь вывод один – азиатский базис;
- Percrocutidae: здесь тоже вывод согласно экскурсионным изысканиям по датировке и местоположению находок. Самые ранние географически обрамляют южные побережья Чёрного и Средиземного морей (возможно, наследники азиатских мигрантов). Затем - типовая форма Percrocuta, ареал которой в среднем миоцене охватывал Малую Азию, южные регионы Средиземноморья, восточную часть северной Африки и Монголию - северный Китай (в обход Гималаев, не иначе). А вот Dinocrocuta уже укрепила границы – доминантное циркумсредиземноморское распространение с регистрацией остатков в позднем миоцене Сивалика и некислая конкурентная позиция в гильдии хищников в териокомплексах верхнего миоцена северо-восточной Азии. Практически одновременное появление этой зверюги в Европе и северном Китае при отсутствии образцов Percrocuta, хоть несколько способных свидетельствовать об эволюционных преобразованиях в сторону терминальной формы, также подталкивает на мысль о загадочном эволюционном азиатском «штабе». А изначально зудит, конечно, и происхождение перкрокутовых от форм, схожих с Africanictis - уж очень африканские стеноплезикты интересны, т. к. появляются просто ниоткуда и в никуда растворяются;
- Barbourofelidae; по ним отписывался, однако здесь несколько сложнее вследствие наличия и последующего прохореза таксонов «дочерних» эволюционных узлов. Наиболее ранние находки, представляющие хищников уже чётко очерченной группы, известны из Африки. Однако скомпилировать сколько-нибудь стройную филогению среди африканских «барбов» очень затруднительно при наличии аргументов хронологических, геологических, фаунистических и прочих факторов, которые напрочь «разрывают» всяческие предположения о возможной морфологической связи между таксонами. Каково филогенетическое положение Afrosmilus и Prosansanosmilus по отношению к друг другу или примитивному Ginsburgsmilus? Откуда вообще взялся Syrtosmilus или европейский Sansanosmilus, появившийся практически одновременно с конгруэнтной азиатской формой? Ну, хотя бы отрадно, что североамериканский кластер общепризнанно является наследником азиатских родов;
- И, ясень пень, Felidae… Proailurus – не прямой потомок европейских стеноплезиктовых, а лишь «осколок» обширнейшей мегапопуляции археофелид, охватывавшей всю Голарктику с позднего олигоцена по ранний миоцен. Первое доказывают явные отличия от прогрессивных Stenoplesictidae, второе – наличие образцов фелоидов проайлуринной категории в хронологически схожих отложениях Азии и Северной Америки, в сумме позволяя предположить очередной экспорт эволюционно продвинутых форм в периферийные регионы из Азии. Затем – Pseudaelurus, на протяжении миоцена как внедрявшего, так и подпитывавшего различные филогенетические линии кошачьих. Хотя и стоит признать, что находки коническозубого Styriofelis turnauensis из Западной Европы датируются чуть раньше, чем у образца подобного морфотипа в раннем миоцене Китая (Sihong), однако тогда стоит соразмерить Азию с Европой и насколько последняя изрыта, так что это не столь существенный аргумент.
Должен он быть, некий кипящий «котёл», постоянно брызгающий эволюционными инновациями.
Только где она - фелоидная Шамбала, мля)))?…
Спасибо всем, кто до конца осилил мои излияния.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#481 27 August 2017 20:02:40
- shish02
- Гость
Re: Эволюция кошкообразных (Feliformia)
С родством Nimravidae и Nandiniidae как-то не клеится. В палеоцене и эоцене Африка изолирована и очень далеко от Азии и от Южной Европы (Южная Европа, по сути, представляла гряду более или менее крупных островов). Даже в олигоцене Африка все еще далеко от Евразии. Если в олигоцене сухопутный путь из Юго-Восточной Азии в Европу просматривается четко (вдоль северной границы Паратетиса; хотя и здесь, кажется, не без разрывов), то Африка остается изолированной. Хотя от Испании ее отделяет совсем небольшое расстояние, северо-восточная Аравия все еще под водой и связи с Азией в восточном направлении нет. По большому счету коридоры для миграций из Африки в Азию и наоборот появятся только в раннем миоцене.
Нандинии уж очень архаичны. Не знаю известны ли их останки за пределами Африки.
#482 28 August 2017 21:37:27
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5935
Re: Эволюция кошкообразных (Feliformia)
Андрей, всё, что знаю по фоссильным находкам Nandinia – находка какого-то зуба из кенийской локации (Tugen Lukeino), с датировкой 6,1 млн, упомянутого в фолианте Верделина (Werdelin 2010. Cenozoic Mammals of Africa). Фото и описания образца нет, он найден Б. Сенут из национального музея естественной истории Франции. Перелопатил доступные работы этой дамы и, единственное, что обнаружил в «поминаньях» - то, что по её предположению, некоторые посткраниальные фрагменты задних конечностей обезьян вполне могут принадлежать циветтам, а вовсе не приматам.
Затем вчера заполночь и сегодня полдня штудировал следующие буквари:
1. Wiig, 1985. Relationship of Nandinia binotata (Gray) to the Superfamily Feloidea (Mammalia, Carnivora)
2. Hunt, 1987. Evolution of the Aeluroid Camivora: Significance of Auditory Structure in the Nimravid Cat Dinictis
3. Hunt, 1989. Evolution of the Aeluroid Carnivora: Significance of the Ventral Promontorial Process of the Petrosal, and the Origin of Basicranial Patterns in the Living Families
4. Joeckel et al., 2010. The auditory region and nasal cavity of Oligocene Nimravidae (Mammalia: Carnivora)
5. Wible & Spaulding, 2013. On the cranial osteology of the african palm civet, Nandinia binotata (gray, 1830) (Mammalia, Carnivora, Feliformia)
Ну, и попутно всякое «по диагонали»... Хант, безусловно, монстр! Исключительно скрупулёзен, фундаментален и последователен, таких сейчас мало, кто пишет. Единственное упущение в том, что он рассматривает барбурофелиевых как филогенетических потомков нимравид и на основе этого изначальный эволюционный вектор выстраивает, а потому в его выводах не исключены заблуждения. Но тут не стоит забывать, что с того времени почти три десятилетия минуло.
Выяснил лично для себя следующее (не вдаюсь в специфические особенности):
- «хрящевая» основа слухового пузыря у хищников не всегда является примитивной характеристикой, так как зависимость оссификации элементов буллы от экологических условий не выявлена вообще начиная с 70-х гг, хотя наблюдается её генезис в эволюционно прогрессивных таксонах. Но, во всяком случае, топология буллы Nandinia действительно примитивна и подтверждает монофилию айлуроидов, эволюционировавших отдельно от арктоидных и каноидных морфотипов:
- в то же время окостенение буллы или её отдельных частей с воззрением, что хрящ как соединительная ткань не обязательно выглядит примитивизмом, очень полезны при онтогенетическом сравнении, анализе и предположениях о филогенетическом родстве семейств Carnivora – это очень даже наблюдательно в онтогенезе гиенид и герпестидовых;
- у одного образца Nandinia binotata (AMNH-M 51488) каудальная часть буллы полностью окостеневшая, хотя это ювенильная особь с наличием молочных зубов. Причём морфологически она совершенно ничем не отличается от серии образцов нандиний с хрящевыми капсулами (где-то у Ханта исследование и анализ проводились). И это в очередной раз подтверждает, что «хрящевидность» не является безапелляционно центральной в вопросе примитивности (это на заметку Артёму Александровичу). Основа, в первую очередь – сама форма слухового пузыря и соотношения его элементов.
- ввиду того, что Nandinia является единственным современным представителем Carnivora с хрящеморфной булллой, это позволяет хотя бы примерно сравнить её с архаичными представителями нимравидовых (Dinictis). Конструктивно пузыри у них даже очень схожи, однако наличествует ряд расхождений (наиболее существенное в связи буллы с петрозалью, формирующей срединную стенку, демонстрируемое у нимравид);
- морфология слуховых капсул очень вариативна в современных группах хищных млекопитающих, а гиалинность (хрящевая основа) её элементов прослеживается у ювенильных особей гиен и герпестид как предковое состояние. Это может быть объяснено предположением, что они являлись онтогенетически пластичными в период эволюционной истории Carnivora на уровне дивергенции на аркто-каноидов и фелоидов. Сложно выразился, согласен))) Суть в том, что морфология эоценовых фелоидов была ещё диффузорна, а именно в олигоцене и происходит их референтность. Разделение на Nimravidae/Nandinidae и на другие формы, почему-то эволюционно устойчивые в решении проблем эффективности и чувствительности слуховых качеств среди представителей Feloidea до сих пор наблюдаются в структуризации слуховых регионов или отдельных компонентов ювенилов. Очевидно, тут целый спектр адаптационных приспособлений сыграл в плане «низложения» архаичных Aeluroidea, енто ж эволюция…
- есть ещё такой нюанс, как кондилоидное экзоципитальное отверстие, отделённое от заднего рваного и объединённого у большинства современных хищных (подспудно подозреваю, что это именно со структурой слуховой буллы связано), но это так… во всяком случае, не сейчас;
Под занавес:
Андрей, прекрасно осведомлён о палеарктических фаунообменах и в курсе, что Африка внедрилась в Евразию лишь в позднем олигоцене. Тут довольно весомое подтверждение – отсутствие находок эоцен-олигоценовых нимравидин в африканских регионах. Но здесь двоякое предположение о генезисе Nandinidae - или реликт какой-то азиатской группы, либо же наследник опять-таки азиатских примитивных хищников, впоследствии адаптировавшихся к экологическим условиям. По сути, хронологической корреляции нандиниевых с нимравидами нет, но всё же…
Спекулятивно опять-таки, выползает гипотеза о превентивной сердцевине эволюционной азиатской Альфы фелоидов. Ведь и нимравиды нам известны уже в стадии очень даже диагностических таксонов. Но всё же где-то было это «ядрышко», где произошло разделение на нимравидо- и виверррообразных; этот «котёл», где впоследствии выпестовывались все генералистичные «опоки», из которых в миоцене формировались мангусты, иктитерии, гиены, перкрокутовые, барбурофелиды и кошки…
Большое спасибо Андрею Владиславовичу за вышележащий пост, стимулировавший вскрытие «белого пятна» в приватной компетенции по фелидоморфным хищникам.
Ну, и самое главное - без «ощупывания» и личного лицезрения материала рассуждать на эту тему не стоит, так что данный вопрос в своих личных изысканиях закрываю.
Всё… ((((
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#483 29 August 2017 16:10:09
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Извините, что прерываю вашу интересную беседу, очень интересно, а то я по кошкообразным отстал вообще.
Игорь, ты как специалист по фелиформиям можешь определить хоть примерно что это за вид или хотя бы род? http://forum.zoologist.ru/viewtopic.php … 50#p488750 Снято якобы в Гирском лесу. Я бы определил ее как представителя рода генетт, но это Гирский лес. Хотя возможно, конечно, что перепутали названия и это вовсе не Гир.
Также я тебюе оставил сообщение вот здесь - http://forum.zoologist.ru/viewtopic.php … 18#p488518
Отредактировано Амфицион (29 August 2017 17:12:37)
#484 29 August 2017 17:28:44
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Из-за различий в морфологии и образе жизни оба подсемейства мадагаскарских хищников ранее определялись как минимум в два разных семейства: мунго (Galidiinae) относили к мангустовым, а малагасийских циветт (Euplerinae) к виверровым, в то время как таксономическая принадлежность фоссы оставалась спорной. Из-за некоторых характерных черт её иногда даже относили к семейству кошачьих, хотя наиболее часто всё же к одной из первых двух групп.
Генетические исследования посредством сравнения ДНК выявили неожиданный результат, что мадагаскарские хищники образуют монофилетическую группу, то есть происходят от общего предка. После этого их начали выделять в отдельное семейство Eupleridae. Филогенетические родственные отношения внутри этого таксона ещё окончательно не выяснены и возможно, что мунго являются парафилетической группой.
Наиболее близкими родственниками мадагаскарских хищников являются мангусты. Вероятно, мадагаскарские хищники произошли от мангустообразного предка, пересекшего в позднем олигоцене или в раннем миоцене (от 20 до 30 миллионов лет назад) Мозамбикский пролив. Таким образом, по сравнению с другими мадагаскарскими группами млекопитающих они являются относительно молодым таксоном, который несмотря на это за короткое время смог занять разные экологические ниши.
Литература
D. E. Wilson & D. M. Reeder: Mammal Species of the World. Johns Hopkins University Press, 2005, ISBN 0801882214
A. Yoder und J. Flynn:Origin of Malagasy Carnivora. In: S. M. Goodman und J. Benstead (Hrsg.): The Natural History of Madagascar. PDF
Источник — «http://ru.wikipedia.org/wiki/%D0%9C%D0%B0%D0%B4%D0%B0%D0%B3%D0%B0%D1%81%D0%BA%D0%B0
#485 29 August 2017 18:22:45
- shish02
- Гость
Re: Эволюция кошкообразных (Feliformia)
Амфицион :
Извините, что прерываю вашу интересную беседу, очень интересно, а то я по кошкообразным отстал вообще.
Игорь, ты как специалист по фелиформиям можешь определить хоть примерно что это за вид или хотя бы род? http://forum.zoologist.ru/viewtopic.php … 50#p488750 Снято якобы в Гирском лесу. Я бы определил ее как представителя рода генетт, но это Гирский лес. Хотя возможно, конечно, что перепутали названия и это вовсе не Гир.
Также я тебюе оставил сообщение вот здесь - http://forum.zoologist.ru/viewtopic.php … 18#p488518
Возможно, это Малая цивета (Viverricula indica). Обитает в том числе и в тех краях.
#486 29 August 2017 19:23:15
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Да, это она!!! Посмотрел несколько фоток.
#487 30 August 2017 16:09:07
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5935
Re: Эволюция кошкообразных (Feliformia)
Так точно, это расса.
С виверрами всё очень непросто. Помимо того, что в них подчас таксоны в различные подгруппы перебрасывают, так и новое семейство могут выделить))).
Хотя по мне, так для бОльшего порядка среди современных можно ещё и подсемейство Genettinae обозначить с типовым родом и африканским линзангом (Poiana), попутно убрав Genetta piscivora и возродив род Osbornictis. А вот в какое подсемейство определить ископаемых Kichechia и Legetetia - фиг его знает, но они достаточно от современных родов отличаются для обособления в собственную группу. Помимо всяческих генетических анализов здесь серьёзная ревизия нужна с подробным рассмотрением фоссильных образцов. Ввиду архаичных черт виверры самая сложная группа из кошкообразных.
Сейчас набросал филограмму:
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#488 31 August 2017 12:07:23
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Тут ты сблизил нандиний с нимравидами. Я как-то писал об этом.
Но почему перкрокуты ближе к кошачьим, чем к другим группам?
Твою работу по перкрокутам я читал не до конца, попал в больничку, а файл остался в Грузии, кажется.
#489 31 August 2017 15:27:59
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5935
Re: Эволюция кошкообразных (Feliformia)
Тады, брат, ты прочёл только первый десяток страниц, не больше, посему и спрашиваешь. Не воспринь за менторство или бахвальство, но это эссе сам Семёнов одобрил (могу даже скан его письма предоставить), так что в данном вопросе не хочу терять время на комментарии. Если нужно, перешлю заново, делов-то… Тут больше интересует систематическая позиция барбурофелид, потому что инфы с описанием образцов, способных просветить по филогении, ничтожно мало.
По больничке искренне сожалею, в своё время в госпиталях тоже чуть ли не по полгода иногда валялся, раны залечивал, ну его на фиг, даже вспоминать не хочется... Надеюсь, что сейчас ты «в седле».
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#490 22 September 2017 18:24:01
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Перепощу сюда, так как это самый посещаемый раздел и все спецы сконцентрированы тут
Еще, у хищных хищнический зуб верхней челюсти это четвертый премоляр, а у кошек третий. Как так?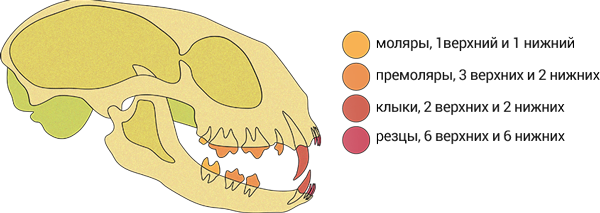
Ведь хищнические зубы хищных гомологичны. Или кошка потеряла четвертый премоляр верхней челюсти? Но как такое возможно, если почти не действующий верхний моляр имеется.
Или это потому что первый премоляр у кошек утрачен? А за клыком идет второй премоляр?
Отредактировано Амфицион (22 September 2017 18:26:09)
#491 23 September 2017 22:13:34
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5935
Re: Эволюция кошкообразных (Feliformia)
Разумеется, в верхней челюсти за клыком - второй премоляр. В окклюзии он не участвует. Из личных фото:
ЗИН, P.leo, № 5723

ЗИН, P.onca, № 5739
К слову, Р2 (или выраженные альвеолы таковых) у всех есть. Ну, во всяком случае в тех черепах, что сам изучал - тигры, львы, леопарды, ягуары, снежные барсы, пумы, гепарды, ягуарунди, оцелоты и пр. Рысей не смотрел. Кстати, по размеру у большинства они практически соразмерны М1 (хотя у тигров и ягуаров последние выглядят всё же больше, но это личное наблюдение). Интересно, что у одного образца Neofelis nebulosa он выглядел как обломанная спичка, находясь на одинаковом расстоянии между клыком и Р3.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#492 23 September 2017 22:45:59
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Благодарствую.
#493 24 September 2017 15:13:43
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Пумы ведь вымерли в Северной Америке, а потом вновь ее заселили из Южной. Они значит прошли через ьутылочное горлышко как и гепарды? А гепарды почему прошли через бутлочное горлышко?
#494 25 September 2017 14:54:19
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5935
Re: Эволюция кошкообразных (Feliformia)
Для начала давай разберёмся, что ты сам понимаешь под термином "бутылочное горлышко"?
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#495 26 September 2017 16:45:41
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Блин вечра же писал ответ, куда он делся?
Сorvin :
Для начала давай разберёмся, что ты сам понимаешь под термином "бутылочное горлышко"?
Сокращение генофонда, который потом восстанавливается.
Игорь, ты не в курсе чем структура клана гиен отличается от волчьей стаи? Я на эту тему инфы нигде не находил.
И еще, в клане гиен самка даже с самым низким положением стоит выше любых самцов. Но ведь самки могут быть мельче самцов. Как тогда эти самки доминируют над самцами? Размеры ведь перекрываются у самцов и самок.
К клану могут присоединяться другие гиены, но это самцы. А самки не присоединяются я так понимаю?
В некоторых источниках пишут что клан гиен это семейная группа, в других об этом не указывается. Стаи волков это семейные группы к которым могут присоединяться одинокие волки. А клан гиен состоит насколько я помню из нескольких семейных групп. В этом отличсие клана гиен от стаи волков?
И получается что все самки всех семейных групп более доминантны по отношению у самцам разных групп клана, так? И как они избегают инбридинга если самка может спариваться с кем попало?
Отредактировано Амфицион (26 September 2017 17:05:18)
#496 26 September 2017 17:33:27

Re: Эволюция кошкообразных (Feliformia)
Амфицион :
Блин вечра же писал ответ, куда он делся?
Сorvin :
Для начала давай разберёмся, что ты сам понимаешь под термином "бутылочное горлышко"?
Наверное здесь http://forum.zoologist.ru/viewtopic.php?id=1325 пост #355 ![]()
Отредактировано Banderass (26 September 2017 17:34:29)
Я вижу тонкие миры, там демоны, парят силуэтами эфимерными... ©
Неактивен
#497 26 September 2017 17:40:41
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Черт, не туда запостил. Спасибо.
#498 26 September 2017 17:56:00
Re: Эволюция кошкообразных (Feliformia)
Амфицион :
Блин вечра же писал ответ, куда он делся?
Сorvin :
Для начала давай разберёмся, что ты сам понимаешь под термином "бутылочное горлышко"?
Сокращение генофонда, который потом восстанавливается.
Артем, в любой популяции могут быть цикличные/периодические изменения/сокращения численности ведущие в свою очередь к сокращению (я бы сказал обеднению или снижению разнообразия) того самого генофонда. Но все это не критично. Критичен порог, при котором остается одна, с большой долей вероятности родственная, изолированная малочисленная популяция. В таком случае вид страдает от инбредной депрессии - обеднение генофонда - накапливание генетических мутаций - низкая жизнеспособность потомства. Если популяция смогла продержаться какой-либо промежуток времени чтоб восстановить численность - считай, она прошла это самое "горлышко". Не смогла - канула в лету. Это как я понимаю, может кто поправит.
Я вижу тонкие миры, там демоны, парят силуэтами эфимерными... ©
Неактивен
#499 26 September 2017 19:02:06
- Crazy Zoologist
- Гость
Re: Эволюция кошкообразных (Feliformia)
Ну да, верно. Гепарды через него прошли. Был у них такой период. Почему не знаю, поэтому и спрашиваю. У пум по-ходу тоже, так как современные пумы происходят из Анд.
#500 26 September 2017 20:51:27
Re: Эволюция кошкообразных (Feliformia)
Амфицион :
Ну да, верно. Гепарды через него прошли. Был у них такой период. Почему не знаю, поэтому и спрашиваю. У пум по-ходу тоже, так как современные пумы происходят из Анд.
Артем, у пум все же генетическое разнообразие определенно есть и разница популяций заметна даже наглядно, чего про гепардов не скажешь. Так что вовсе не факт, что они тоже прошли "бг".
Отредактировано Banderass (26 September 2017 20:52:22)
Я вижу тонкие миры, там демоны, парят силуэтами эфимерными... ©
Неактивен