Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#1301 22 April 2017 12:58:04
- Crazy Zoologist
- Гость
Re: Медведи
Генетики вычислили общего предка белых и бурых медведей
http://www.membrana.ru/particle/13206
#1302 25 May 2017 20:41:16
- Crazy Zoologist
- Гость
Re: Медведи
#1303 13 June 2017 20:39:25
- Crazy Zoologist
- Гость
Re: Медведи
Справка об эволюции современных медведей
Семейство медвежьих (Ursidae), как и другие семейства подотряда собакообразных (Caniformia) эволюционировали из небольших древолазающих хищных млекопитающих, принадлежащих семейству миасовых, или миацид (Miacidae) надсемейства миакоидов (Miacoidea) приблизительно 25 миллионов лет назад (Herrero 1999). Все восемь современных видов медведей происходят от общего предка – урсава, или урсавуса (Ursavus), который появился в ходе эволюции в субтропической зоне на территории современной Европы в течении миоценовой эпохи более чем 20 миллионов лет назад (Craighead 2000). В течении этой эпохи урсавоидные медведи постепенно увеличились в размерах, а их зубы, изначально харакерные для типичных хищных млекопитающих, видоизменились и преобрели черты, свойственные для всеядных животных, такие, как например более плоская и менее режущая коронка предкоренных и в особенности коренных зубов (Ward and Kynaston 1995), что характерно для всеядных животных с большим акцентом в сторону растительноядной пищи.
От урсава развилось три линии современных медвежьих. Согласно молекулярным данным, раньше всего от обещего ствола современных представителей семейства отделилась ветвь, ведущая к большой панде (Ailuropoda melanoleuca). Недавний филогенетический анализ выявил, что, из всех восьми ныне существующих видов медведей, Ailuropoda melanoleuca является наиболее древним (Gittleman 1999, Waits et al. 1999).
Эволюционная ветвь большой панды в лице её предка агриарктоса (Agriarctos) отделилась от общего эволюционного ствола с урсавом примерно 20 миллионов лет назад (Ward and Kynaston 1995). Молекуоярные исследования также показывают, что следующей ветсью, отделившемуся от общего эволюционного ствола, ведущему к современным видам, была ветвь очковых, или андских медведей (Tremarctos ornatus) (Waits et al. 1999), которая отделилась от рода Ursavus примерно 14 миллионов лет назад (Ward and Kynaston 1995) и в настоящее время очковый медведь является единственным видом медвежьих в Южной Америке (Nowak, 1991, Stirling 1993, Craighead 2000).
Прямым прародителем третьей эволюционной линии современных медвежьих, которые представляют собой подсемейство Ursinae (так называемые настоящие медведи) является Protoursus, который эволюционировал из Ursavus elemensis между 12 и 10 миллионами лет назад. Первым, так называемым настоящим медведем (представителем подсемейства урсин) был Ursus minimus, который появился примерно 5 миллионов лет назад. Его непосредственным предком был Protoursus. чьим прямым предков является Protoursus (Ward and Kynaston 1995). Анализ митохондриальной ДНК показал, что медведь-губач является наиболее древним современным представителем подсемейства урсин (Waits et al. 1999, Gittleman 1999).
#1304 29 June 2017 21:54:56
- Crazy Zoologist
- Гость
Re: Медведи
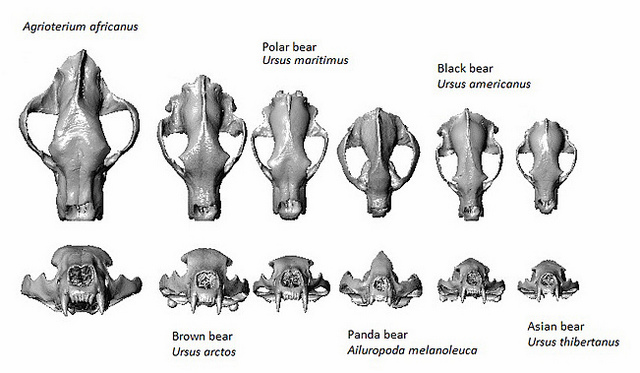
#1305 15 September 2017 21:56:06
- Crazy Zoologist
- Гость
Re: Медведи
Tremarctos floridanus - ископаемый вид семейства медвежьих (Ursidae), распространённый в Северной Америке в эпоху позднего плиоцена (поздний бланкий) - позднего плейстоцена (ранчолабреаний). Возможно дожил до голоцена (Kurten, Anderson, 1980). Ископаемые останки были найдены во Флориде (Devil's Den), юго-востоке США и в Мексике.
Часто флоридского тремарктоса называют флоридским, или американским пещерным медведем в связи со схожей экологической нишей с настоящими пещерными медведями (подрод Spelaearctos), однако эти медведи мало родственны между собой. Ближайшим же родственником T. floridanus является ныне живущий очковый медведь (Tremarctos ornatus), принадлежащий к тому же роду (Tremarctos). T. floridanus вместе с очковым медведем, а также ископаемыми родами Arctodus, Arctotherium и Plionarctos обычно выделяют в подсемейство Tremarctinae и трибу Tremarctini. Барышников же (Барышников, 2007) выделяет этих медведей в трибу Arctotheriini в рамках подсемейства Ursinae. В данном случае Tremartini и Arctotheriini являются синонимами и принципиальная разница заключается в том, что одни учёные склонны выделять тремарктин/арктотериин в отдельное подсемейство, а другие включают его в центральное подсемейство медведей - Ursinae.
T. floridanus был заметно крупнее своего современного родственника и достигал размеров довольно крупного бурого медведя (Ursus arctos). Кондилобазальная длина черепа у самцов T. floridanus составляла около 32 см, у самок - 25,7 см. Половой диморфизм был выражен очень сильно: самцы примерно в полтора раза были крупнее самок.
Лицевой отдел черепа укорочен, с заметным уступом в области лба. Парокципитальный отросток отклонён назад. Нижний край нижнечелюстной кости выпуклый. Щёчные зубы T. floridanus крупнее и относительно шире, чем у очкового медведя. Для флоридского тремарктоса как и для всего подсемейства тремаркторин (Tremarctinae) [fhfrnthyj кулисное смыкание коренных зубов.
Для флоридского тремарктоса предполагают образ жизни очень схожий с таковым у пещерного медведя (Ursus spelaeus). Пещерный медведь представлял собой одну из крайних специализаций среди медведей к растительноядному образу жизни и как и копытные животные был пастбищным животным. Вероятно, что флоридский тремарктос занимал очень похожу с пещерными медведями нишу. Его ближайший родственник очковый медведь также потребляет преимущественно растительные корма, несмотря на то, что его коренные зубы имеют кулисное захождение, что больше характерно для плотоядных, нежели растительноядных животных. Однако строение коренных зубов явственно говорит о преимущественно растительноядной диете медведей рода Tremarctos. Как уже отмечалось выше, щёчные зубы флоридского тремарктоса были ещё более уплощены, чем у очкового, тем самым жевательная поверхность коронки зубов была ещё большей, чем у последнего. Скорее всего это связано с тем, что флоридский тремарктос в большей степени, нежели его современный родственник, потреблял травянистые растения и больше времени проводил на земле. Очковый же медведь отлично лазает по деревьям и проводит на них довольно много времени, заодно питаясь там и отдыхая. Вероятно, что флоридский пещерный медведь использовал пещеры в качестве убежищ, однако нет явных свидетельств его залегания в пещеры, как это делал европейский пещерный медведь. Возможно, что в виду более мягкого климата Флориды (даже в эпоху плейстоцена) американский экологический аналог европейского пещерного медведя в спячку не впадал.
Систематика
Класс:
Mammalia (млекопитающие, или звери)
Подкласс:
Theria (живородящие млекопитающие, или настоящие звери)
Инфракласс:
Eutheria (плацентарные, или высшие звери)
Надотряд:
Laurasiatheria (лавразиатерии)
Отряд:
Carnivora (хищные)
Подотряд:
Caniformia (собакообразные, или псообразные)
Инфраотряд:
Arctoidea (арктоидные, или арктоиды)
Надсемейство:
Ursoidea (медведеобразные)
Семейство:
Ursidae (медвежьи)
Подсемейство:
Tremartinae или Ursinae (тремарктины, или короткомордые медведи или урсины (в зависимости от взгядов на систематику))
Триба:
Tremarctini/Arctotheriini (короткомордые медведи, тремарктины, или арктотериины)
Род:
Tremarctos (тремарктосы, или очковые медведи)
Вид:
Tremarctos floridanus (флоридский тремарктос, американский пещерный медведь, флоридский пещерный медведь, или флоридский очковый медведь)
#1306 15 September 2017 21:59:13
- Crazy Zoologist
- Гость
Re: Медведи
Этрусский медведь
Общее представление
Этрусский медведь (Ursus (Euarctos) etruscus) - ископаемый вид медведей, обитавший с позднего плиоцена (средний виллафранк) по ранний плейстоцен (поздний виллафранк) плейстоцен в Европе, Азии, а также в Северной Африке. Внешне этрусский медведь несколько напоминал бурого медведя (Ursus arctos) и судя по всему является его предком.
Эволюция и систематика
Этрусский медведь является крупнейшим представителелем подрода Euarctos, куда входят также современные барибалы (Ursus americanus) и гималайские медведи (Ursus thibetanus), последнего, впрочем, некоторые авторы помещают в подрод Selenarctos.
Предком этрусского медведя являлся наиболее мелкий примитивный представитель подрода Euarctos - Ursus (Euarctos) minimus, с которым он сохраняет ряд архаичных черт. В Европе этрусский медведь впервые появляется в позднем плиоцене, примерно 2,5 миллиона лет назад (средний виллафранк) (Mazza, Rustioni, 1992). Более ранние находки данного вида не подтверждены.
В эволюции вида выделяют две стадии. Этрусские медведи ранней стадии обнаружены в Испании, Франции, Болгарии, Греции и Украине. В Азии медведи эти стадии известны из Таджикистана. Имеется также ископаемый материал в виде щёчных зубов, сходных с ранним Ursus etruscus, из среднего виллафранка Марроко, Северная Африка (Geraads, 1997).
Этрусский медведь вероятно был довольно теплолюбивым видом, так как обширный ареал распространения этого вида ограничен почти исключительно южными широтами Палеарктики. Этрусский медведь найден в Средиземноморью, Центральной Азии, Кавказе и в Китае. Наиболее северная находка этрусского медведя известна из местонахождения Тегелен в Нидерландах.
Морфология
Этрусский медведь - самый крупный представитель подрода Euarctos, превосходящий размерами американских и азиатских чёрных медведей и достигающий размеров не очень крупного бурого медведя, на которого он был похож внешне.
Лицевой отдел этрусского медведя широкий и более удлинён, нежели у других представителей подрода Euarctos. Он лишь немногим короче мозгового и составляет примерно 90% от длины последнего. Мозговая коробка слабо расширена в заднем отделе. Кондилобазальная длина черепа самцов этрусского медведя превышает 34 см.
Носовые кости удлинённые, умеренно узкие, впереди почти доходят до заднего края верхних клыков. Носовая апертура небольшая и округлая. Подглазничное отверстие маленькое, лежит над задней частью коронки первого верхнего моляра (M1). Лоб широкий. У некоторых особей уплощённый, у других - выпуклый и слегка приподнят в области глазниц. Глазницы небольшие, округлые. Заглазничные отростки крупные, отонутые вниз. Височное сжатие хорошо выражено. Сагиттальный гребень сильный, особенно ярко выражен у самцов. Костное нёбо широкое. Задненёбная вырезка клиновидной формы. Хоаны узкие. Слуховые барабаны уплощённые. Затылочные мыщелки довольно маленькие (Барышников, 2007). Голова этрусского медведя при жизни очевидно больше напоминала голову бурого медведя, нежели других представителей подрода Euarctos, так как краниометрически этрусский медведь ближе к бурому медведю, чем к чёрным медведям и любым другим из подрода Euarctos.
Клыки сравнительно слабые. Присуствует полный набор передних премоляров P1-3/p1-3. Премоляры маленькие, разделены широкими диастемами. Первые премоляры (P1/p1) не увеличены или же незначительно крупнее остальных передних премоляров. Размеры щёчных зубов превышают таковые у остальных представителей подрода Euarctos.
Верхний хищнический зуб (P4) имеет большой внутренний выступ. Талон на втором верхнем моляре (M2) относительно длинный. Метаконид нижнего моляра (m1) одиночный или двойной. Длина первых нижних моляров (m1) составляет примерно 20,9-24,7 мм, длина верхних моляров - 23,22 мм. Половой диморфизм хорошо выражен. Имеются значительные различия в величине черепа и развитости сагиттальных гребней между самцами и самками (Барышников, 2007).
Подвиды
Барышников (2007) принимает четыре подвида этрусского медведя:
Ursus etruscus etruscus.
Диагноз: передние премоляры мельче, а задние крупнее, чем у Ursus etruscus saintvallierensis. Второй моляр (M2) с относительно широким талоном, первый нижний моляр (m1) с двойным метаконидом, второй нижний моляр (m2) с длинным талонидом.
Распространение и возраст: Европа (Оливоля, Верхнее Вальдарно, Петрафита); ранний плейстоцен (поздний виллафранк).
Ursus etruscus saintvallierensis.
Диагноз: зубы меньше, чем у Ursus etrusscus verescagini. Второй верхний моляр (M2) с узким талоном, первый нижний моляр (m1) с двойным метаконидом, второй нижний моляр (m2) с коротким талонидом.
Распространение и возраст: Франция (Saint-Vallier) и, вероятно, Испания (La Pueblo de Valverde); поздний плиоцен (средний виллафранк).
Ursus etruscus verescagini.
Диагноз: зубы крупные. Первые верхние моляры (M1 и M2) относительно широкие, особенно M1. Нижний хищнический зуб (m1) с одиночным или двойным метаконидом, второй нижний моляр (m2) с умеренно длинным талонидом.
Распространение и возраст: Таджикистан (Куруксай, Оби-Гарм, Тутак); поздний плиоцен (средний виллафранк).
Ursus etruscus vekuai.
Диагноз: зубы крупнее, чем у других подвидов, особенно увеличен второй верхний моляр (M2). Первый нижний моляр (m1) с двойным метаконидом. Верхние щёчные зубы по средним значениям промеров превосходят таковые у подвидов из Европы и Таджикистана. Крупными размерами зубов, особенно M2, Ursus etruscus vekuai напоминает бурого медведя, однако у него присуствуют все передние премоляры.
Распространение и возраст: Грузия (Дманиси); ранний плейстоцен (поздний виллафранк) (Барышников, 2007).
Образ жизни
Сходство в размерах, зубной морфологии и пропорциях метаподиальных костей этрусского медведя с бурым предполагает, что этрусский медведь по образу жизни был похож на последнего. Он обитал в разнообразных ландшафтах, преимущественно лесных. Приуроченность этрусского медведя к южным областям, вероятно, указывает на связь этого вида с угодьями богатыми растительными кормами. Это обстоятельство, по-видимому, не позволило ему проникнуть через Берингийский мост в Северную Америку (Барышников, 2007). Судя по всему, этрусский медведь был менее склонен к хищничеству, нежели его бурый родственник. Не исключено, что этрусский медведь, как и бурый, залегал в спячку для зимнего сна, однако его южное распространение и отсуствие скопления костей в пещерах позволяют предположить, что его спячка не была обязательной (Барышников, 2007).
Систематика
Класс:
Mammalia (млекопитающие, или звери)
Подкласс:
Theria (живородящие млекопитающие, или настоящие звери)
Инфракласс:
Eutheria (плацентарные, или высшие звери)
Надотряд:
Laurasiatheria (лавразиатерии)
Отряд:
Carnivora (хищные)
Подотряд:
Caniformia (собакообразные, или псообразные)
Инфраотряд:
Arctoidea (арктоидные, или арктоиды)
Надсемейство:
Ursoidea (медведеобразные)
Семейство:
Ursidae (медвежьи)
Подсемейство:
Ursinae (урсины)
Триба:
Ursini (урсины)
Род:
Ursus (медведи)
Подрод:
Euarctos (эуарктосы, или чёрные медведи)
Вид:
Ursus (Euarctos) etruscus (этрусский медведь)
Подвиды:
Ursus etruscus etruscus
Ursus etruscus saintvallierensis
Ursus etruscus verescagini
Ursus etruscus vekuai
#1307 20 September 2017 17:58:49
- Crazy Zoologist
- Гость
Re: Медведи
Можно ли медведя назвать мезохищником? Гиперхищник - 100% хищник. Мехохищник - средний. А как называются хищные млекопитающие, животная пища которых составляет мизерный процент? Гипохищники?
#1308 20 September 2017 18:57:24
- Алекс
- Сан Саныч - Модератор

- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
Амфицион :
Можно ли медведя назвать мезохищником? Гиперхищник - 100% хищник. Мехохищник - средний. А как называются хищные млекопитающие, животная пища которых составляет мизерный процент? Гипохищники?
веганхищник !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1309 20 September 2017 19:53:18
- ZaecЪ
- Любознательный

- Откуда: Киев
- Зарегистрирован: 12 July 2017
- Сообщений: 158
Re: Медведи
Алекс :
Амфицион :
Можно ли медведя назвать мезохищником? Гиперхищник - 100% хищник. Мехохищник - средний. А как называются хищные млекопитающие, животная пища которых составляет мизерный процент? Гипохищники?
веганхищник !
колеблющийся веган ;-) Посидит месяц-другой на корешках да не удержится и хапанёт колбаски
Неактивен
#1310 20 September 2017 19:56:19
- ZaecЪ
- Любознательный
- Откуда: Киев
- Зарегистрирован: 12 July 2017
- Сообщений: 158
Re: Медведи
Амфицион :
Можно ли медведя назвать мезохищником? Гиперхищник - 100% хищник. Мехохищник - средний. А как называются хищные млекопитающие, животная пища которых составляет мизерный процент? Гипохищники?
Гиперхищник ещё "облигатный хищник", а гипохищник так и будет (хотя мне больше нравится "факультативный хищник" — питается выпускниками факультета :-D ). И медведи как раз относятся к гипохищникам, а не к мезо-
Отредактировано ZaecЪ (20 September 2017 20:10:26)
Неактивен
#1311 20 September 2017 20:35:54
- Crazy Zoologist
- Гость
Re: Медведи
ZaecЪ :
Амфицион :
Можно ли медведя назвать мезохищником? Гиперхищник - 100% хищник. Мехохищник - средний. А как называются хищные млекопитающие, животная пища которых составляет мизерный процент? Гипохищники?
Гиперхищник ещё "облигатный хищник", а гипохищник так и будет (хотя мне больше нравится "факультативный хищник" — питается выпускниками факультета :-D ). И медведи как раз относятся к гипохищникам, а не к мезо-
Факультативный хищник может очень часто питаться мясом. Факультативный хищник это нехищное изначально животное, которое стало хищным. Например неандерталец - факультативный хищник. Он много мяса ел.
#1312 20 September 2017 23:11:04
- ZaecЪ
- Любознательный
- Откуда: Киев
- Зарегистрирован: 12 July 2017
- Сообщений: 158
Re: Медведи
Насчёт"очень часто" сомневаюсь, это как облигатный и факультативный аэроб — первые живут в строго кислородной среде и немедленно погибают без его доступа, а вторые обычно анаэробные, но выживающие и слабо размножающиеся в присутствии кислорода. Так и с питанием мясом…
Неактивен
#1313 21 September 2017 21:12:27
- Crazy Zoologist
- Гость
Re: Медведи
Почему сомневаетесь? Неандертальцы и мы - факультативные хищники. Некоторые народы до сих пор едят много мяса и оно составляет основу их рациона.
#1314 21 September 2017 22:12:43
- Crazy Zoologist
- Гость
Re: Медведи
Затылочный гребень это производное затылочного бугра?
Барышников пишет, что геологическая история губача практически неизвестна, однако также пишет, что род известен из позднего плиоцена. Как это понимать?
Монография Барышникова в виде книги вышла в 2007 году. А саму работу когда он написал? Если даю ссылку на монографию Барышникова, то какой год указывать? 2007?
Отредактировано Амфицион (21 September 2017 22:56:59)
#1315 22 September 2017 16:28:42
- Crazy Zoologist
- Гость
Re: Медведи
Губач (Melursus ursinus) – один из восьми современных представителей семейства медвежьих (Ursidae). Это единственный современный представитель рода Melursus.
Внешний вид губача довольно своеобразен. Удлинная лохматая шерсть (это самый лохматый вид срези медведей), особенно на плечах. Окраска одноцветная, чёрная, нередко с примесью бурого и серого оттенков. Отмечались также коричневые и красноватые (рыжеватые) особи. Передняя часть морды грязновато-белая (Барышников, 2007). Телосложение массивное с мощными конечностями, крупными лапами и огромными когтями (на передних лапах они намного длиннее, чем на задних). Наряду с малайским медведем, или бируангом (Helarctos malayanus) у губача самые крупные когти относительно своего размера среди всех медведей, по крайней мере современных.
Различают два подвида губача – континентальный Melursus ursinus ursinus и цейлонский – Melursus ursinus inornatus. Губачи с острова Цейлон (Шри-Ланка) заметно мельче континентальных.
Диплоидное число хромосом – 74 (2n = 74, NFa = 84) (Барышников, 2007).
Голова с удлинённой лицевой частью. Ушные раковины длинные, подвижные, дистально расширяющиеся, в основании трубчатые, усиленные в основании вертикальными хрящевыми гребнями. Ринариум (носовое зеркальце; безволосая область носа вокруг ноздрей) без фильтрума на верхней губе и без медиального желобка (Барышников, 2007). Щелевидные ноздри способны произвольно смыкаться, что предохраняет дыхательные пути от попадания в них пыли и насекомых (Pocock, 1941; Барышников, 2007). Губы почти голые, очень подвижные, имеют способность сильно вытягиваться вперёд. Язык длинный. Строение губ, языка и ринариума связано с питанием муравьями и термитами. Кисть с одиночной наружной карпальной (запястной) подушечкой и маленькой внутренней, или только с участком голой кожи, обозначающим её (Барышников, 2007).
Кондилобазальная длина черепа у самцов 263-320 мм, у самок – 256-301 мм. Череп длинный относительно его ширины. Лицевая часть черепа умуренно длинная. Передний край глазницы лежит над М2 (втором верхнем моляре). Костное нёбо сводчатое в области моляров, которые редуцированы и располагаются впереди так, что постдентальная часть нёба не короче Р4-М2. Слуховой пузырь уплощнный, его ширина с костной трубкой нарудного слухового прохода значительно меньше, чем длина ряда верхних щёчных зубов. Нижняя челюсть с длинным симфизным отделом. Первые верхние резцы отсуствуют. При окклюзии (смыкании челюстей) нет отчётливого заклыкового пространства, которое имеется у всех современных представителей медвежьих, кроме рода Ailuropoda (к которому относится большая панда, или бамбуковй медведь (Ailuropoda melanoleuca).
Зубная система губача весьма своеобразна. Нижние резцы при окклюзии могут перекрывать верхние. Оба первых верхних резца утрачены (это единственный случай в отряде хищных), что, наряду с подвижными губами, способными вытягиваться в трубочку, способствует действуию ротовой полости губача как «пылесоса». Клыки относительно небольшие. Ширина верхнего клыка варьирует у самцов от 12,6-ых до 15 мм.
Передние премоляры не уменьшены относительно размеров моляров. Вторые и третьи, как верхние, так и нижние премоляры (Р2-3/р2-3) относительно крупные и разделены диастемами (промежутком между зубами). Третий верхний (P3) и третий нижний премоляр (р3) могут иметь два корня (Р3 чаще всего с двумя корнями). Верхний хищнический зуб Р4 небольшой, с крупным метаконом и хорошо развитой внутренней долей; протокон часто с добавочным зубцом.
М1 прямоугольный, с небольшим парастилем и более крупным метастилем. Метаконуль по высоте не уступает паракону, есть чёткий мезокон. Хорошо выражен переднелингвальный цингулюм. М2 уже, чем М1, его талон слабо дифференцирован от тригона. Оба верхних моляра относительно уже, чем в роде Helarctos (к которому относится малайский медведь).
Нижний премоляр р4 заметно выше, чем параконид м1, с небольшим дистальным зубцом. Нижний хищнический зуб м1 шире в области талонида, чем в области тригонида. Метаконид имеет вид крупного одиночного бугорка. Энтоконид сдвинут к заднему краю зуба, талоидный бассейн не выражен.
Нижний моляр м2 прямоугольный, его талонид значительно короче, чем тригонид, и чаще всего более узкий. Метаконид выше протоконида, иногда различим бугорок параконида. Коронка м3 овальная со стороны жевательной поверхности, с маленьким внутренним полем, не разделена на тригонид и талонид.
Согласно Г. Ф. Барышникову (Барышников, 2007) длина тела составляет 140-180 см, длина хвоста 10-12,5 см. По другим данным, длина тела составляет 140-190 см ( "Bear Anatomy and Physiology" in Brown, Gary (1993) The Great Bear Almanac, Lyons & Burford, Publishers; "Sloth Bear". The Animal Files. Retrieved 18 April 2011; "Sloth Bear". Arktofile.net. Retrieved 18 April 2011; "San Diego Zoo's Animal Bytes: Sloth Bear". Sandiegozoo.org. Retrieved 18 April 2011).
Высота в плечах согласно Г. Ф. Барышникову – 61-91,5 см. По другим данным - 61-90 см (Blomstrom, 2000; International Association for Bear Research and Management (IBA), September 22, 1999; Sanderson, 1972; Ward and Kynaston, 1995). По другим данным 60-90 см ( "Bear Anatomy and Physiology" in Brown, Gary (1993) The Great Bear Almanac, Lyons & Burford, Publishers; "Sloth Bear". The Animal Files. Retrieved 18 April 2011; "Sloth Bear". Arktofile.net. Retrieved 18 April 2011; "San Diego Zoo's Animal Bytes: Sloth Bear". Sandiegozoo.org. Retrieved 18 April 2011). (Самцы примерно на 30-40% крупнее/тяжелее самок (Барышников, 2007; (Blomstrom, 2000; International Association for Bear Research and Management (IBA), September 22, 1999; Sanderson, 1972; Ward and Kynaston, 1995).
Масса тела самцов варьирует от 80-ти до 140-ка кг, у самок от 55-ти до 95-ти кг (Nowak, 1991; Барышников, 2007). По другим данным, масса тела самцов составляет 80-192 кг, со средним весом 130 кг, а самок – 55-124 кг ( "Bear Anatomy and Physiology" in Brown, Gary (1993) The Great Bear Almanac, Lyons & Burford, Publishers; "Sloth Bear". The Animal Files. Retrieved 18 April 2011; "Sloth Bear". Arktofile.net. Retrieved 18 April 2011; "San Diego Zoo's Animal Bytes: Sloth Bear". Sandiegozoo.org. Retrieved 18 April 2011).
В настоящее время геологическая история рода Melursus почти неизвестна. Морфологически губач хорошо обособлен от других представителей триби Ursini, куда он входит. Наряду с признаками высокой специализации, связанной с энтомофагией (питанием насекомыми) данный вид имеет ряд примитивных (архаичных) особенностей – передние премоляры с двумя корнями, отсуствие при окклюзии свободного пространства над премолярами, отсуствие первых верхних резцов.
В связи с этим, губач выглядит архаичнее не только других представителей Ursini, но и медведей трибы Arctotheriini. Анализ ядерной ДНК помещает род Melursus как сестринский таксон рода Helarctos (к которому принадлежит малайский медведь). Комбинированный анализ ядерной и митохондриальной ДНК определяет базальное положение губача в трибе Ursini (Li et al.,2004a; Барышников, 2007). Скорее всего род Melursus рано отделился от предкового ствола. Удлинение лицевого отдела происходило у него параллельно таковому роду Ursus (Барышников, 2007).
Палеонтологические документы крайне скудны. Лидеккер (Lydekker, 1884) описал вид Ursus theobaldi по почти полному, но сильно повреждённому ископаемому черепу без зубов из района Кангры (Kangra) В Индии (Himachai Pradesh). Размеры черепа крупные, кондилобазальная длина черепа около 335 мм (расчитана по рисунку: Lydekker, 1884, Pl XXVIII). По строению костного нёба Ursus theobaldi похож на Melursus ursinus.
Пилгрим (Pilgrim, 1932 : 52) первым отнёс вымерший вид к роду Melursus, рассматривая его в качестве непосредственного предка современного губача. Судя по альвеолам, щёчные зубы его были крупнее, чем у современного вида. Есть также незначительные отличия в форме слухового пузыря и нижнечелюстной ямки (Colbert et al., 1935; Edbrink, 1953; Барышников, 2007), но эти отличия не превышают уровня подвидовых. Стратиграфическое положение находки из Кангры соотвествует, вероятнее всего, позднему плиоцену (верхнему сивалику).
Судя по маленьким размерам щёчных зубов и сводчатому строению костного нёба к Melursus ursinus (то есть к тому же виду, что и современный губач) принадлежит череп из пещеры Чандрапал (Chandrapalle) на юге Индии, по которому описан вид Ursus footei (Prassad, Yadagiri, 1986; Барышников, 2007). Хотя у последнего нёбо более узкое, а морда несколько короче, чем у губача, эти различи могут объясняться возрастной изменчивостью. Непосредственные останки Melursus отмечены в других плейстоценовых местонахождениях В Индии (Billasurgam Cave) (Prassad? 1996; Барышников, 2007).
Губач рапространён в Индии, Шри-Ланке (Цейлон), Бангладеше, Непале и Бутане. Раньше он встречался практически по всей Индии, однако сегодня его ареал сократился, но он всё ещё широко распространён в этой стране.
Населяют губачи главным образом влажные и сухие тропические и субтропические леса, особенно в районах с выходом к скалам. Также их можно встретить на травянистых равнинах. Предпочитает небольшие высоты, обычно не поднимаясь выше предгорий. Умеет хорошо лазать по деревьям, однако делает он это редко и не забирается на них при опасности. Несмотря на свои относительно небольшие размеры это очень сильное и смелое животное, которое иногда может дать отпор даже тигру.
Активен в любое время суток, но главным образом ночью. Самки с детёнышами и молодые особи активнее в дневные часы, а взрослые самцы – в ночные и сумечечные. Это связано, вероятно с тем, что самки больше подвержены нападению крупных хищников, которые обычно активны в ночные часы и будучи активны днём они минимализируют нападение с их стороны. Но в целом, как уже было сказано выше, это ночные животные, которые проводят день в густых зарослях или пещерах, особенно в тех местах, где поблизости есть вода. В спячку губачи не впадают, однако в сезон дождей их активность понижается.
Как и все медведи, губач ведёт одиночный образ жизни, за исключением самок с детёнышами и периода размножения. Наблюдение за одним взрослым самцом в национальном парке Читван, ранее называемым Королевским национальным парком Читван (Royal Chitwan National Park) в Непале показало, что он в течение почти двух лет по площади, не превышающей 10 кв. км. Минимальная плотность популяции в том же районе составляла 0,1-ых особи один кв. км (Laurie, Seidensticker, 1977; Барышников, 2007).
Имеется значительное перекрывание индивидуальных участков взрослых особей одного пола и взрослых и полувзрослых особей обоих полов. Зарегистрировано перемещение самцов в муссонный период с аллювиальных пойменных равнин в более возвышенные саловые леса. Самки не имеют сезонных перемещений (Joshi et al., 1995; Барышников, 2007).
Обоняние развито превосходно, однако зрение и слух оставляют желать лучшего. Из-за этого нередко происходят сталкновения этих медведей с людьми, которые заканчиваются весьма плачевно – губачи иногда уродуют людей, нанося им страшные раны своими огромными когтями. Во время сталкновения с соперниками или другими крупными хищниками губачи рычат, ревут, издают пронзительные крики и визг. Чтобы напугать врага губачи, что характерно для представителей семейства медвежьих встают на задние лапы, чтобы казаться больше.
Большую часть года основу рациона губачей составляют термиты – до 50% от общего обзёма пищи. Поедаются также и другие насекомые (например муравьи) и их личинки, мёд, яйца, падаль, трава, цветы и различные плоды (Барышников, 2007), а также листья. Благодаря своим мощным когтям губач разрушает термитник, складывает губы и язык в трубочку, сдувает пыль, а затем буквально всасывает их. Любит он также разорять гнёзда пчёл. Длинным языком он может доставать насекомых из труднодоступных мест. Ноздри губача, как уже отмечалось выше, могут смыкаться, тем самым защищая дыхательные пути от попадания туда пыли или насекомых. Возможно, при удобном случае губач может убить мелкое или раненое животное, но таких зарегистрированных случаев пока неизвестно, хотя чисто физически он способен убивать не только мелких животных. Процент той или иной пищи в рационе губачей варьирует в зависимости от времени года. С марта по июнь процент плодов в рационе губачей возрастает и может составлять до 50% его диеты (Joshi et al., 1997; Ward and Kynaston, 1995; Sanderson, 1972; Барышников, 2007).
Брачный период в Индии приходится в основном на июнь, в Шри-Ланке медведи могут спариваться в течение большей части года. Во время спаривания губачи ведут себя очень шумно (Blomstrom, 2000). Новорожденные появляются на свет главным образом с сентября по январь. Беременность длится 6-7 месяцев. Самка рождает, как правило, одного или двух детёнышей, реже трёх. Рождаются медвежата обычно в пещере или земляном убежище, которое покидают примерно через 1,5-2 месяца. Детёныши открывают глаза примерно примерно в трёхнедельном возрасте. Подросших детёнышей самка часто переносит на спине. Удивительно, что во время опасности детёныши даже будучи на дереве предпочитают слезть и забраться на спину матери, которая спасается бегством (Laurie, Seidensticker, 1981; Joshi et al., 1997; Барышников, 2007). Медвежата остаются с матерью до двух-трёх лет. К этому моменту они почти полностью вырастают (Ward and Kynasaton, 1995; Blomstrom, 2000; Барышников, 2007). Продолжительность жизни в природе толком неизвестна. В неволе губач доживал до 40-ка лет (Ward and Kynaston, 1995; Барышников, 2007).
Естественными врагами губача являются тигр (прежде всего) и леопард. Верояятно и дхоли, или красные волки представляют для него определённую опасность, но достоверных случаев убийства губача стаей дхолей на данный момент неизвестно. Жертвой крупного леопарда может стать самка губача, особенно некрупная, ну и естественно молодняк. Однако даже самка губача представляет собой весьма сильное животное и леопарды обычно не связываются с ними. На взрослых самцов губачей леопарды не нападают. Единственный хищник, который представлет для них опасность это тигр, но и он довольно редко нападает на взрослых самцов.
#1316 27 September 2017 14:28:26
- Crazy Zoologist
- Гость
Re: Медведи
У Барышникова даты неточные. Кто в итоге старше индарктос или агриотерий? Агриотерий дожил до плиоцена, индарктос вымер раньше, но согласно вике появился тоже раньше. Хотелось бы знать точные даты ихз существования. В цифрах или с указанием классических ярусов из геохронологической шкалы.
Заранее спасибо.
#1317 04 October 2017 13:08:41
- Wolf
- Без пяти минут зоолог

- Зарегистрирован: 13 January 2009
- Сообщений: 2583
Re: Медведи
Индарктос старше и был период, когда они жили в одно и то же время и кажется, даже в одних и тех же местообитаниях. Как делили не знаю.
Per aspera ad astra
Неактивен
#1318 07 October 2017 20:34:37
- Crazy Zoologist
- Гость
Re: Медведи
Точных датировок по индарктосу не нашел. Он появляетс в позднем, а может и в среднем миоцене. Вымирает судя по всему тоже в течение миоцена, в отличие от агриотерия.
#1319 11 October 2017 11:42:25
- Crazy Zoologist
- Гость
Re: Медведи
У меня закралось такое ощущение, что крупные медведи типа индарктоса, агриотерия, арктодуса и т.п. вышли в открытые места благодаря своим размером. Барышников пишет, что медведи преобретя крупные размеры смоглиосвоить открытые пространства. Но потом крупные длинноногие медведи вымирают. Не связано ли это с приобретением социальности у крупных кошачьих?
См. подпись:-)
#1320 14 October 2017 13:06:29
- Crazy Zoologist
- Гость
Re: Медведи
Странно как-то эволюционируют медведи современного типа. Сначала примитивные урсавусы, потом уже специализированные виды такие как индарктос и т.п.
Отредактировано Амфицион (14 October 2017 13:09:14)
#1321 14 October 2017 17:14:46
- Crazy Zoologist
- Гость
Re: Медведи
Я вот подумал, не Тибет ли является родиной бурого медведя? Именно там обитает подвид, сохранивший в зрелом возрасте ювенильную окраску. У Барышникова на эту тему ничего не нашел, там в основном находки из России и сопредельных стран.
#1322 14 October 2017 18:51:43
- Crazy Zoologist
- Гость
Re: Медведи
https://auction.catawiki.com/kavels/833 … m-standing вот тут тоже говорится о ключице пещерного медведя. Пишут что ее длина 33 см. Не слишком ли дофига?
#1323 15 October 2017 14:32:25
- Crazy Zoologist
- Гость
Re: Медведи
Амфицион :
У меня закралось такое ощущение, что крупные медведи типа индарктоса, агриотерия, арктодуса и т.п. вышли в открытые места благодаря своим размером. Барышников пишет, что медведи преобретя крупные размеры смоглиосвоить открытые пространства. Но потом крупные длинноногие медведи вымирают. Не связано ли это с приобретением социальности у крупных кошачьих?
См. подпись:-)
Алекс твое мнение интересует, так как ты любишь саблезубых и много о них знаешь. Мнения остальных тоже интересует. Обожаю мишек, больших да малых.
#1324 15 October 2017 19:29:04
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
Амфицион :
Амфицион :
У меня закралось такое ощущение, что крупные медведи типа индарктоса, агриотерия, арктодуса и т.п. вышли в открытые места благодаря своим размером. Барышников пишет, что медведи преобретя крупные размеры смоглиосвоить открытые пространства. Но потом крупные длинноногие медведи вымирают. Не связано ли это с приобретением социальности у крупных кошачьих?
См. подпись:-)
Алекс твое мнение интересует, так как ты любишь саблезубых и много о них знаешь. Мнения остальных тоже интересует. Обожаю мишек, больших да малых.
Кто от чего вымирал меня мало интересует ,все равно это в 90 % предположения и фантазии !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1325 17 October 2017 02:06:54
- Crazy Zoologist
- Гость
Re: Медведи
Совершенно верно. В палентологии вообще многие вещи лишь предположения. Иногда это интригует, иногда раздражает. Ну это как по мне. Просто меня уже несколько "подташнивает" от того, что все все валят на изменение климата и истребление человеком животных. Индарктос и агриотерий так или иначе схожи, но один перешагнул черту миоцена, другой нет. И тот который перешагнул (агритерий) вымирает в плиоцене, в то время когда вроде никаких катаклизЬмов не было. Если были, то пусть меня поправят. Я буду только рад. Он заселил в том числе и Америку, но в итоге вымирает, но затем приходят ведмеди другой группы (тремерктины (подсемейство) или арктотериини (триба) в зависимости от взглядов тех или иных ученых на систематику ведмедей. И потом тоже вымирают в конце плейстоцена, как и многие дргие хищники (более хищные чем ведмеди), хотя многие виды относящиеся к мегафауне к их потенциальной добыче) доживают до голоцена.
Немного позволю себе отклониться от темы: львы в Америке вымирают еще до того как вымирает одна из их основных жертв - бизонов. Которых было просто "дохрена" даже в гораздо более поздние времена, когда белые осваивали запад. К тому же эти бизоны стали более уязвмы для хищников, так как стали меньше и рога их стали менее эффективны для противостояния.
Все это с одной стороны меня привлекает: предположения, основанные на логике и т.п. С дркгой стороны как бы отбивает охоту к изучению ископаемых видов.