Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#1201 19 March 2017 10:41:11
- Алекс
- Сан Саныч - Модератор

- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
Амфицион :
Но в таблицах не указан диаметр кости умки. У кого толще кости непоняятно.
 В таблицах промеров толщины нет ,наглядно вроде особой разницы нет .
В таблицах промеров толщины нет ,наглядно вроде особой разницы нет .
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1202 19 March 2017 10:42:46
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
Амфицион :
Алекс :
http://www.archaeology.org/news/5381-170316-polar-bear-skull http://savepic.ru/13244092m.png Ну и в чем гигантизм ,если этот череп немного больше 40см ?
Ну у белого ведь череп меньше чему бурого, для белого наверное это много.
{kind=link}
В таблицах у белого современного череп до 441 мм . А здесь немного больше 40 см ,да еще необычно узкий !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1203 19 March 2017 13:09:55
- Crazy Zoologist
- Гость
Re: Медведи
Алекс, если кости примерно одинаковые, а голова у брого больше начит по максимуму он все же крупнее?
#1204 19 March 2017 13:15:49
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
Амфицион :
Алекс, если кости примерно одинаковые, а голова у брого больше начит по максимуму он все же крупнее?
Сомневаюсь , что 3-4 см разницы черепа в длине ,что то решают в массе , все индивидуально .
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1205 19 March 2017 15:08:18
- Crazy Zoologist
- Гость
Re: Медведи
Не только в длине, но и в ширине и высоте + передняя часть туловища не так сильно развита.
#1206 19 March 2017 15:17:38
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
Амфицион :
Не только в длине, но и в ширине и высоте + передняя часть туловища не так сильно развита.
Ну судя по фото плечевых костей в сравнении ,дистальные и проксимальные концы больше у белого . Да и в общем белый сложен по другому !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1207 19 March 2017 15:31:18
- Crazy Zoologist
- Гость
Re: Медведи
,это же не гонка в размерах ,как у динозавров
Почему нет? К тому же тираннус был крупным, потом измельчал.
Получается что огромный умка теперь оотится на мелких нерп, более крупных тюленей берет намного реже, про моржей даже молодняк я уже молчу.
Зачем емук такие размеры тогда?
Отредактировано Амфицион (19 March 2017 15:31:37)
#1208 19 March 2017 16:17:59
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
Амфицион :
,это же не гонка в размерах ,как у динозавров
Почему нет? К тому же тираннус был крупным, потом измельчал.
Получается что огромный умка теперь оотится на мелких нерп, более крупных тюленей берет намного реже, про моржей даже молодняк я уже молчу.
Зачем емук такие размеры тогда?
Ничего в тиранусе нет сверх крупного ,судя по чучелам умок в музеях середины 20 века ,умки были не меньше тирануса ! Измельчали умки по двум причинам ,крупных выбили в прошлом столетии и сокращение территорий льдов !Скоро и этих не останется ,только в зоопарках !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1209 19 March 2017 17:28:52
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Медведи
From "Scaling of mammalian long bones: small and large mammals compared" by Per Christiansen, ratio of averaged least circumference and humerus length values:
Ursus arctos (n=8)= 0.314
Ursus maritamus (n=6)= 0.325
Urus middendorfi (n=2)= 0.358 Вот нашел показатель наименьшей окружности диафиза плечевой кости по Христиансену .
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#1210 19 March 2017 18:32:10
- Crazy Zoologist
- Гость
Re: Медведи
Алекс, в среднем кто крупнее кадьяк или умка?
#1211 19 March 2017 23:43:30
- Crazy Zoologist
- Гость
Re: Медведи
Читал, что у белого медведя кости плотнее, чем у бурого, и это объяснялось адаптацией к водному образу жизни. Кто может логически это объяснить? Чтобы легче нырять было? С друкгой стороны жир который держит на плаву, одно другому противоречит.
#1212 20 March 2017 00:03:06
- Crazy Zoologist
- Гость
Re: Медведи
Арктотерий.

#1213 20 March 2017 00:37:47
- Crazy Zoologist
- Гость
Re: Медведи

#1214 21 March 2017 15:29:27
- Crazy Zoologist
- Гость
Re: Медведи
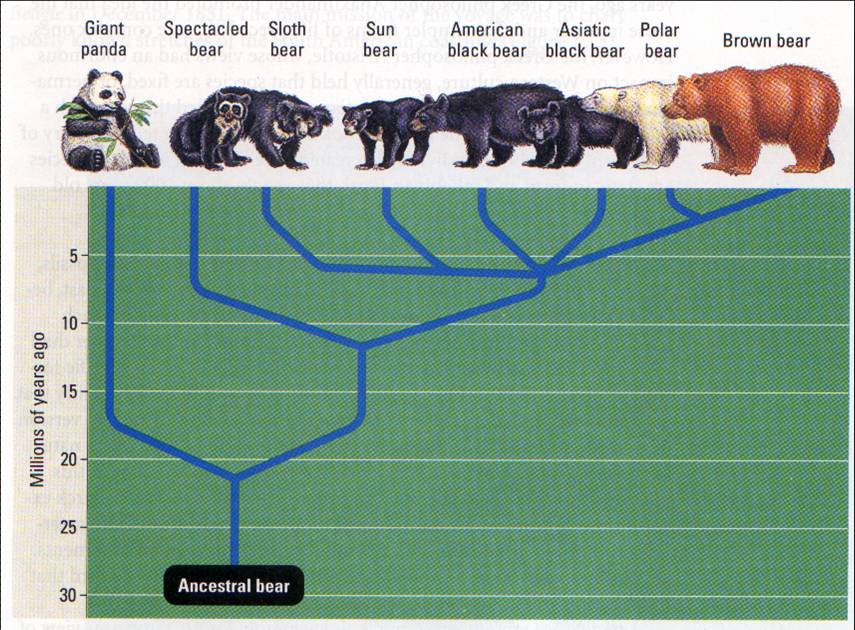
#1215 21 March 2017 15:32:56
- Crazy Zoologist
- Гость
Re: Медведи

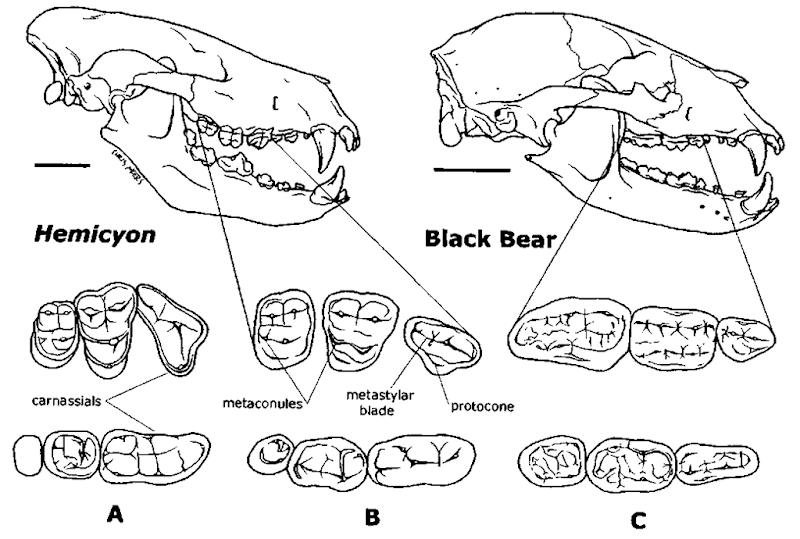
Какова ориентировочная максимальная масса тела для гемициона?
#1216 21 March 2017 15:35:42
- Crazy Zoologist
- Гость
Re: Медведи
В вике очень разная инфа, Легендре и Ротт измеряли массу тела и у них то меньше 100 кг получалось то около 600 кг.
#1217 21 March 2017 23:56:42
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Медведи
АртёмСаныч, на уровне твоей компетенции и в силу возраста (что подразумевает умудрённость и опыт) оперировать инфой из Вики напросто непрофессионально, уж прости.
В моём воспалённом воображении хемиционовые выглядят очень даже обособленной группой. Пока не готов достойно аргументировать, всё весьма ещё аморфно, да и пока интересуют другие хищники, но завязывается эмбрион гипотетического предположения, что хемиционы являются на самом деле аберрантной филогенетической линией арктоидных хищников, отличающейся от собственно медведей.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#1218 22 March 2017 00:14:34
- Crazy Zoologist
- Гость
Re: Медведи
Но там ссылка на источник - Legendre & Roth.
хемиционы являются на самом деле аберрантной филогенетической линией арктоидных хищников, отличающейся от собственно медведей.
Я тоже к этому склоняюсь, но Барышников объединяет их с медведями.
#1219 22 March 2017 20:29:03
- Crazy Zoologist
- Гость
Re: Медведи
В связи с жизнью во влажном тропическом климате ископаемые останки малайского медведя крайне скудны. Большая редкость находок бируанга в континентальных местах может также объясняться давлением со стороны гималайского медведя, который в плейстоцене вытеснял своего более мелкого родственника. Наибольшее число ископаемых останков малайских медведей обнаружено на островах Борнео, Суматра и Ява, которые датируются поздним плейстоценом (Medway, 1964; Vos, 1983). В настоящее время континентальные находки бируанга известны лишь из Вьетнама.
В китайской провинции Шанси был обнаружен фрагмент правой нижнечелюстной кости с первым моляром, по которому Зданский (Zdansky, 1927) описал вид Ursus cf. boekhi. Точное стратиграфическое положение данной находки неизвестно, предположительно она относится к плиоцену. Тениус (Thenius, 1947a) обратил внимание, что тело нижнечелюстно кости заметно повышается к симфизу, как у Helarctos и на основании этого выделил китайскую находку в вид Ursus (Helarctos) sinomalayanus. Единственный сохранившийся зуб m1 (первый моляр) несколько крупнее, чем у современного малайского медведя; его длина составляет 20,7.
Метаконид одиночный, расположен немного позади протоконида. Эрдбринк (Erdbrink, 1953) предполагал, что метаконид был двойным, но его передний бугорок слился с ребром, отходящим от вершины протоконида. Гипоконид большой, энтоконид отчётливый, имеется энтоконулид. По середине талона проходит продольный желобок, не замкнутый сзади, как это часто имеет место у H. malayanus (Барышников, 2007). Впоследствии на основании развития эмалевых зубчиков впереди метаконида m1 эта находка была отнесена к Ursus minimus (Morlo, Kundrat, 2001).
Другой вид, U. praemalayanus, был основан на ископаемом правом M2, вероятно купленным в китайской аптеке в Сингапуре или Гонконге (Koenigswald, 1935). Автор описания отмечал сходство нового вида с малайским медведем, а Эрдбринк (Erdbrink, 1953) прямо писал, что не нашёл между ними существенных различий. Барышников для этой находки условно принимает подвидовой статус.
Родственные связи малайского медведя с другими представителями семейства остаются невыясненными. Стандартное генетическое расстояние Нея между малайским и бурым медведем (Ursus arctos) оказалось меньше (0,025), чем таковое между малайским медведем и гималайским медведями (0,037). Данные молекулярной биологии противоречивы. На основе полных последовательностей митохондриальной ДНК бируанга сближают либо с кладой Ursus arctos / maritimus (Talbot, Shields, 1996a), либо с американским чёрным медведем (Ursus americanus) (Zhang, Ryder, 1993a). Исследования ядерной ДНК продемонстрировали генетическую близость малайского медведя с губачом. Комбинированный же анализ ядерной и митохондриальной ДНК, напротив, удаляет его от губача (Li et al., 2004). Родственные связи с губачом поддерживаются морфологическими данными, а также случаями гибридизации этих видов в неволе (Van Gelder, 1977).
#1220 22 March 2017 22:13:11
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Медведи
Коллеги, вот сколько насчёт «ведмедко» бьёмся, а постоянно возникают вопросы, подчас граничащие с некоторой неосведомлённостью, как это не прискробно признать.
Потратил пару-тройку вечеров на небольшой ликбез, представляю:
Исследования по систематике медвежьих в первую очередь базируются на изучении морфологии. Морфологически семейство представляет хорошо очерченную группу: половой диморфизм выражен только в размерах; конечности мощные, укороченные и стопоходящие; череп крупный и удлинённый, с выраженными сагиттальным и затылочным гребнями и сильными скуловыми дугами. Слуховые капсулы небольшие, камера образована преимущественно эктотимпаником. Нижняя челюсть массивная и длинная, с широким сочленовным отростком.
Клыки крупные. Верхний хищнический зуб Р4 меньше M1, паракон высокий, протокон крупный, парастиль отсутствует. Карнассиальное лезвие метастиля преобразовано в округлый метакон. Нижний хищнический зуб р4 удлинённый, иногда наличествует короткий талонид. Коренные зубы прямоугольные, давящего типа, с дополнительными вершинками.
Посткраниальные кости у медвежьих сходной морфологии, характерно увеличение количества крестцовых позвонков, передние конечности используются для манипулирования гораздо больше, чем у других хищников.
Как представляется, первые урсоиды появились в конце среднего эоцена. Ранние формы были, наверное, небольшими всеядными древолазающими животными, а увеличение размеров обусловлено наземным образом жизни. В эволюции характерны увеличение массивности черепа и нижнечелюстной кости, а также формирование мощных скуловых дуг, что связано с развитием жевательной мускулатуры.
Основные направления адаптаций не вполне ясны, как проблематичны и этапы дивергенции семейства. Постараюсь осветить несколько популярно в несколько постов:
Семейство Subparictidae
Проблематичная группа, объединяющая мелких арктоидов позднего эоцена - раннего олигоцена (Parictis, Nothocyon, Campylocynodon, Drassonax) с расширенными премолярами, которая некоторыми исследователями принимается как базальная для урсоидов. Другие полагают некоторые таксоны (Parictis, Drassonax) близкими к Amphicynodontinae, но это окончательно не доказано.

Нижнечелюстная ветвь Parictis

Реконструкция Nothocyon. Только, конечно, вряд ли она правильна ввиду явной канидообразности. Скорее они были похожи на злоупотреблявших стероидами куньих

Череп Campylocynodon (DCM № 50-26-6919)
Достигали размеров куницы и, хотя посткраниаль неизвестена, предположительно это были стопоходящие или полустопоходящие длиннохвостые зверьки, ведущие древесный и полудревесный образ жизни.
P. S. Скорее всего, сборная группа. Корректней было бы просто обозначить её как Ursoidea или Arctoidea incertae sedis.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#1221 22 March 2017 22:14:56
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Медведи
Семейство Ursidae
Следующая группа – подсемейство амфицинодонтин (Amphicynodontinae). Это зверушки посерьёзней, так как включают уже генерализованных медвежьих.

Верхний зубной ряд и нижняя челюсть Amphicynodon
Триба Amphicynodontini
Включает Amphicynodon и Pachycynodon - мелких амфицинодонтин из раннего олигоцена Европы размером с куницу, с укороченной лицевой частью черепа и примитивной зубной морфологией.
Триба Amphicticipini
Amphicticeps – относительно специализированные формы из раннего олигоцена Монголии с гипертрофированным развитием мастоидных отростков. Это не присуще другим амфицинодонтинам, но характерно для североамериканских родов Allocyon и
Kolponomos.
Allocyon – схож с колпономом.
Kolponomos приспособлен к обитанию в водной среде, на что помимо краниальных характеристик указывают и элементы зачерепного скелета.
Череп (LACM 131148) и нижняя челюсть (USNM 215070) Kolponomos newportensis

Морда очень ндравится реконструкционно, а вот с посткраниалью не очень согласен, уж слишком медведеподобна (но это отнюдь не критика Р. С. Учителя).

Аналогично у Р. Евсеева. Технически исполнено безупречно, да и краниологически подогнано идеально, но уж явно выражена «каланоморфность».
Безусловно, амфицинодонтины не являлись предками ластоногих, а принадлежность к Ursidae подтверждается наличием слёзной кости, отсутствующией у ластоногих; небольшим задним рваным отверстием (foramen lacerum), которое у ластоногих крупное, а также отличным развитием мастоидной области (этот признак отрицает возможность водного образа жизни).
Особняком в группе состоит такой род, как Adelpharctos из позднего олигоцена Франции. Хотя высокий, заострённый p4 и развитый метаконид на ml сближают его с амфицинодонтинами, конкретно идентифицировать его пока невозможно.
P. S. По-видимому, амфицинодонтины – базальные палеарктические формы плюс «аппендиксная» североамериканская филогенетическая линия (Allocyon и Kolponomos), которая может являться потомком азиатских видов.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#1222 22 March 2017 22:17:58
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Медведи
Подсемейство Hemicyoninae
Размеры от средних до крупных. Череп массивный, с широкими скуловыми дугами и крупными хищническими зубами. Нижнечелюстная кость высокая, с премассетерной ямкой. Главной особенностью группы является пальцехождение, что позволило хемиционинам освоить эконишу псовых и гиенид. С этим связано наличие таких признаков, как удлинение лицевого отдела черепа, укрупнение карнассиалов, редукция внутреннего бугорка на Р4 и метаконида на ml. Несмотря на то, что некоторыми исследователями хемиционины признавались даже как отдельное семейство, их зубная система выглядит вполне урсидной.
Появляются в Европе в позднем олигоцене, в Северной Америке - в раннем миоцене. Центр эволюции – Евразия.
Триба Cephalogalini 
Череп Phoberogale
Cephalogale - включает несколько форм, занимающих базальное положение в подсемействе (например, ввиду отсутствия премассетерной ямки, имеющейся у остальных хемиционин). В пределах группы наличествует ряд эволюционных уровней и направлений специализации. Демонстрируют удлинение хищнических зубов, что позволяет считать их родственными более поздним гиперкарниворным родам.
Триба Hemicyonini
Различаются 3 эволюционные линии. Первая характеризуется усилением режущей специализации зубной системы.
Phoberocyon - крупные курсориальные хищники. Для зубной системы характерна мозаичность примитивных характеристик (рЗ-4 с задними дополнительными вершинками, талонид на ml схож с таковым у Cephalogale) с прогрессивными (метаконид на ml более редуцированный, чем у Hemicyon, р4 крупнее р3 и т.д.).

Реконструкция Plithocyon
Plithocyon – среднеразмерные хищники, Ml-2 прямоугольные, их талоны небольшие. Передние премоляры низкие, упрощённые; р4 без задней дополнительной вершинки; метаконид на ml уменьшен; протоконид и метаконид на m2 одинаковой высоты.
Zaragocyon - из раннего миоцена Испании, обладающий промежуточными признаками между Cephalogale и [i]Hemicyon. Известен по щёчным зубам, по строению и пропорциям моляров сходен с Plithocyon.

Реконструкция Hemicyon (дон Маурицио)
Вторая группа - дробяще-режущей специализации: Parahemicyon и Hemicyon. Вместе с первой одновременно появляются в раннем миоцене Европы. В Северной Америке известны представители только первой линии, которые зарегистрированы немного позднее, чем в Европе.

Челюстные фрагменты Hemicyon sansaniensis
Hemicyon - нижние передние премоляры не уменьшены, задних дополнительных вершинок не имеется, что отличает его от Phoberocyon. Нижние премоляры не редуцированы, как у Plithocyon. Не ревизован.
Третьей линии из среднего и позднего миоцена Европы присуща зубная система дробящего типа.
Dinocyon - наиболее крупные представители группы, массивные звери с мощными зубами и сильными конечностями. Возможно, являлись остеофагами и по образу жизни напоминали крупных гиен.

Зубы и челюсти Dinocyon thenardi
Несмотря на разнообразие пищевых предпочтений хемиционин, их локомоция уступала псовым, кошачьим и гиенидам. После миграции канид хемиционины вымирают, очевидно, не выдержав конкуренции с этими группами.
P. S. Цефалогаловые – гетерогенная группа, дифференцируемая на роды Ginsburgocyon, Phoberogale и собственно Cephalogale. Не исключено, что ранние и примитивные формы этого рода могли дать начало Ursavus, тогда как поздние и более прогрессивные – Hemicyoninae.
Очень интересен вопрос по американским хемиционинам среднего миоцена - Plithocyon barstowensis и Р. ursinus в связи с тем, что Plithocyon известен только из Европы и не регистрируется в Азии. В то же время в раннем миоцене Казахстана и Китая фиксируется Phoberocyon johnhenryi и Р. youngi (MN4), которые моложе европейского Phoberocyon aurelianensis (MN3). Т. е. миграция ранних хемиционин в сторону Нового Света вполне обозначена. И, очевидно, предположения Цю (Qiu, 2003. «Dispersals of Neogene camivorans between Asia and North America») о том, что североамериканские хемициониды принадлежат к отдельному эндемичному таксону (производному от азиатских фобероционов), вовсе не лишено оснований.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#1223 22 March 2017 22:26:52
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Медведи
Подсемейство Ursinae
Появляются в раннем миоцене наряду с хемиционинами. Для этой группы характерно развитие дробяще-давящей функции зубов щёчного ряда, при которой режущая способность пары хищнических зубов снижается, а затем утрачивается. Редукция карнассиалов сопровождалась увеличением площади задних моля¬ров, причём уникальным признаком является формирование длинного талона на М2, что не наблюдается у представителей других семейств Carnivora. Передние премоляры либо увеличиваются и моляризуются (Ailuropodini), либо уменьшаются в размере (Arctotheriini, Ursini).
Триба Ursavini
Ursavus – базальный таксон, от которого, вероятно, произошли все урсины. Известен с миоцена Евразии и Северной Америки.

Реконструкция Ursavus

Череп и нижнечелюстная ветвь Ursavus tedfordi
Урсавусы - короткомордые медведи, у которых Р4 демонстрирует тенденцию к уменьшению в размерах; М1 почти квадратный; на М2 наблюдается прогрессивное удлинение талона; метаконид на ml крупный; m2/3 чуть крупнее, чем ml; параконид на m2 почти не дифференцирован, метаконид выше протоконида, талонид укорочен.
Судя по скелету раннего U. orientalis, он обладал вытянутыми лапами, длинным хвостом и экстерьерно напоминал виверру. Возможно, предпочитал древесный образ жизни. Более поздние и крупные урсавусы уже были более склонны к сухопутности - метаподиальные кости из среднего миоцена Европы сходны с таковыми современного малайского медведя. Возможно, это было стимулировано аридизацией климата и попутно сопровождалось увеличением размеров.
Триба Ailuropodini
Морфобиологическое своеобразие предполагает их длительную эволюцию, в ходе которой зубной аппарат достиг наивысшего среди урсид уровня специализации в обработке растительных кормов.
Ailurarctos - поздний миоцен Китая, зубная морфология которого демонстрирует признаки, общие с Ailuropoda. Различия обусловлены более примитивными зубными признаками у Ailurarctos. Размерно примерно на треть меньше современной большой панды.

Находки ископаемых представителей рода Ailuropoda ограничены Восточной Азией, где из раннего плейстоцена известен ввд A. microta, который был меньше современной формы. Современный вид A. melanoleuca появляется в среднем плейстоцене – крупный подвид А. m. baconi, дентальные признаки которого свидетельствуют, что эта форма была менее специализированной, чем современная.
(?)Kretzoiarctos – средний миоцен (MN8), Испания, изначально идентифицированный как Agriarctos beatrix. Дентальная морфология демонстрирует черты, свидетельствующие о приспособлении к питанию растительной пищей, хотя и не в той степени, что у бамбуковых медведей.
Триба Agriotheriini
Возможно, парафилетичная группа, включающая роды Indarctos и Agriotherium, вопрос о систематическом положении и родственных связях которых остаётся открытым. Хотя Agriotherium и Indarctos во многом сходны, гипотеза о происхождении первого рода от второго спорна – как стратиграфически, так и в остутствии апоморфий, поддерживающих монофилию Indarctos - Agriotherium. Скорее всего, оба рода имеют общего предка.
Реконструкция Indarctos (М. Антон)
Indarctos - поздний миоцен Евразии, Северной Африки и Северной Америки. Короткомордые медведи среднего и крупного размера. Характеризуются крупными карнассиалами наряду с большими дробящими молярами. Наличествуют следующие виды – Indarctos vireti (валлезий Европы), I. arctoides (туролий Европы и Турции), I. atticus (туролий Европы, Африки и Азии), I. punjabiensis (туролий Индии и Китая), демонстрирующий редукцию передних премоляров и высокую географическую изменчивость и I. oregonensis (поздний миоцен С. Америки) – эволюционно самый продвинутый. Наблюдается тенденция к размерному увеличению во времени - если I. arctoides не превышал размеров современного барибала, то I. punjabiensis сравним с бурыми медведями о-ва Кодьяк. Судя по сопутствующим фаунам, I. arctoides был связан с лесными ландшафтами, тогда как I. punjabiensis предпочитал более открытые стации.

Череп Agriotherium

Реконструкция Agriotherium
Agriotherium – поздний миоцен-ранний плиоцен Европы, Азии и Африки; ранний плиоцен Северной Америки. Размеры крупные, передние премоляры более редуцированы, чем у Indarctos, Р4 длиннее M1, с увеличенным парастилем, метаконид на ml увеличен и весьма сдвинут.
Признаётся вид A. sivalense, занимавший обширный ареал с несколькими географическими подвидами, которые ранее описывались под отдельными видовыми названиями. Интересны образцы из Нового Света, архаичной дентальной морфологией указывающие на раннюю миграцию сравнительно примитивных форм из Евразии. Другой вид, A. intermedium, известен из позднего плиоцена Польши и Китая – размерно меньше, чем A. sivalense. Имеется также A. inexpetans из позднего миоцена Китая. Дентально схож с A. intermedium, однако таксоны разделены значительной хронологической лакуной.
P. S. Ursavus - голарктический таксон и, по-видимому, всё же полиморфный. Ранние формы (Ballusia), MN3, известны с раннего миоцена Европы (B. elmensis, B. hareni) и Азии (B. orientalis). Последний известен по типовому образцу и не исключено, что представляет отдельный род.
Несмотря на множество именований Ursavus, наличествуют общие тенденции, позволяющие выделить ранне-среднемиоценового U. brevirhinus (MN4–9), средне-позднемиоценового U. primaevus (MN6–9) и, большей частью, позднемиоценового U. depereti (MN9) с рядом переходных форм, географических рас и т. д. Особняком здесь стоят U. isorei из раннего миоцена Франции, эволюционно схожего с урсавинами начала раннего миоцена (возможно, побочная филогенетическая ветвь); U. ehrenbergi (MN11), известный по голотипу из Греции (Halmyropotamos); U. tedfordi из Китая, размерно и морфологически близкий с U. ehrenbergi; китайский U. sylvestris, размерно также близкий с U. ehrenbergi, но отличающийся редукцией протокона Р4 и большей высотой бугорков щёчных зубов; и североамериканский среднемиоценовый U. pawniensis, морфологически близкий U. рrimaevus Старого Света, но гораздо превышающий его в размерах.
Kretzoiarctos - не исключено, что на самом деле не связан с близким родством с Ailuropodа, представляя собой конвергентную адаптивную линию специализированных форм. Это в какой-то степени подтверждают дентальные остатки Agriarctos из позднего миоцена Венгрии, который в настоящее время синонимизирован с Indarctos cf. vireti. Морфологически ml и m2 у Agriarctos сходны с таковыми у Indarctos vireti, но длина ml заметно меньше, а рЗ и р4 более моляризованы – т. е. Agriarctos более прогрессивен. В то же время он архаичней Kretzoiarctos, хотя и жил несколько позже – возможно, здесь присутствуют несколько параллельных линий, объединённых общим предковым таксоном. Также можно упомянуть дентальные остатки Ursavus depereti из французского местонахождения Соблэ (Soblay), демонстрирующие признаки начальной специализации для обработки жёсткой растительности.
Также требует ревизии род Indarctos – возможно, вплоть до выделения в отдельную трибу Indarctiini. Из европейского валлезия известен I. vireti (MN9), затем его стратиграфически сменяет I. arctoides (MN10-12) – очевидно, это напросто хроновиды и интересен североамериканский I. oregonensis – близкий китайским формам I. lagrelii и I. sinensis, синонимизированными с I. punjabiensis.
В связи с вышеизложенным, возможно, целесообразно возродить род Agriarctos, объединяющий ранние формы урсид Европы, чьи щёчные зубы преобразованы для питания растительными кормами. В него могут входить две формы: ранняя (MN 10) с образцами, в настоящее время отнесёнными к Ursavus depereti - P4-M2 из Соблэ, а также нижнечелюстной фрагмент с m1-m2 из Лузинэ (Luzinay) и типовая Agriarctos gaali (=vighi) из туролия Венгрии (MN 12), квалифицируемая как Indarctos vireti. К слову, Ailurarctos lufengensis тоже сперва упоминали как U. depereti.
Что касается Agriotherium, то ещё стоит упомянуть о потенциально отдельном видовом статусе A. africanum, который морфологией m2 резко отличается от всех других.
А предок Ursini - U. primaevus.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#1224 22 March 2017 22:29:22
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Медведи
И ещё одна группа из данной когорты, триба Arctotheriini.
Эндемичная группа Западного полушария. Характеризуется очень короткой мордой, наличием премассетерной ямки, кроме Plionarctos. Верхний хищнический зуб Р4 не столь укорочен, как у урсин, складки эмали на m2 формируют уникальную для группы Z-образную структуру в талонидном бассейне.
Происхождение Arctotheriini конкретно не выяснено. Дентальные характеристики выглядят производными от Ursavus, хотя уровень развития прогрессивнее благодаря редукции хищнических зубов P4/ml и удлинению талонидного отдела на m2. Данных, свидетельствующих о происхождении арктотеринных медведей в Палерктике нет, но их определённые родственные отношения с Ursavus и Indarctos такое вполне допускают.

Нижнечелюстной фрагмент Plionarctos haroldorum
Plionarctos - наиболее архаичный в трибе Arctotheriini и самый ранний. Среднеразмерный медведь величиной с небольшую особь очкового медведя. От остальных арктотериин отличается плезиоморфными признаками (отсутствие премассетерной ямки, позицией протокона Р4 и т. д.). В роде признаётся два вида, образующих парафилетическую стволовую группу, от которой произошли остальные таксоны. Стратиграфическое положение обеих форм близкое: Р. edensis - поздний миоцен (верхний хемфиллий) и Р. harroldorum - ранний плиоцен (нижний бланкий), морфологические различия заключаются в некоторых дентальных особенностях.
Tremarctos - впервые фиксируется в позднем плиоцене Калифорнии. От Plionarctos отличается наличием премассетерной ямки и немного более длинным талоном на М2. Передние премоляры в нижней челюсти у Т. floridanus иногда отсутствуют, щёчные зубы небольшие. Геологическая история современного Т. ornatus из Юж. Америки неизвестна.

Черепа Tremarctos: A). T. ornatus; B). T. floridanus
Вымерший Т. floridanus обнаружен в раннем плейстоцене Северной Америки, но обычным становится в позднем на юго-востоке США и в Мексике Он крупнее, чем Т. огпаtus, щёчные зубы широкие.

Черепа Arctotherium: A). A. tarijense, B). A. bonariense, C). A. vetustum
Реконструкция Arctotherium
Arctotherium – ископаемые арктотериины Южной Америки. Выделяется два подрода - номинативный, существовавший до голоцена и раннеплейстоценовый Arctoidotherium, иногда позиционируемый как Arctotherium angustidens. Последний отличается уплощением слуховой капсулы (что диагностично для урсид), шириной верхних коренных зубов, укрупнённостью протокона на нижнего премоляра р4. Вообще, морфологическое сходство между обеими формами предполагает их близкое филогенетическое родство.
Предок арктотериин, очевидно, мигрировал в Южную Амери¬ку единожды и все изменения обусловлены адаптивной радиацией.

Черепа Arctodus A). A. simus, B). A. pristinus
Arctodus - крупные медведи с сильно укороченным лицевым отделом черепа, отличающиеся от Arctotherium не столь широкими верхними коренными зубами и уплощёнными слуховыми капсулами. Род включает два вида. A. pristinus более грацилен и с пропорционально меньшими зубами. Биотопически занимал те же территории, что и позднее Tremarctos floridanus. A. simus был массивнее, габаритно превосходивший крупнейшие экземпляры современного бурого медведя (особенно юконская популяция).
P. S. Попутные замечания следующие:
Г. Ф, Барышников отмечает, что Arctotherium tarijense морфологически отличается от других видов и вполне может быть выделен в отдельный род Pseudarctotherium. Буржуи (Trajano & Ferrarezzi, 1995. "A fossil bear from northeastern Brazil, with a phylogenetic analysis of the South American extinct Tremarctinae (Ursidae)") вообще вычленяют А. brasiliensis и А. раmparum в род Pararctotherium, которые Геннадий Фёдорович сводит в синонимию с A. vetustum.
На самом деле, коллеги, здесь Сталинград: на основании всяческих автоа-, синапо- и симплеоморфий Pararctotherium вроде бы признаётся как подрод Arctotherium. Единственное - филогенетическое положение A. (P.) enectum проблематично ввиду скудности материала. С другой стороны, Arctotherium и Pararctotherium могут быть также викариантными таксонами, не так ли?
И вообще, пусть америкосы сами со своими ведмедко разбираются.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#1225 22 March 2017 22:32:37
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Медведи
Теперь уж чисто вэдмэдики…
Урсины (Ursini).
Триба включает медведей с относительно удлинённым лицевым отделом черепа, упрощёнными передними премолярами, удлинением и расширением коренных зубов, постепенно снижающимся тригонидом на ml и развивающимся дополнительными вершинками на этом зубе, а также расширением и удлинением талонида на m2. К трибе принадлежат все современные виды Старого Света и североамериканский Ursus americanus. По сути, корневых роды: Helarctos, Melursus и Ursus.
Губач, Melursus - наиболее архаичный. Специализирован к энтомофаги, нёбо и хоаны расширены, передние премоляры относительно крупные, РЗ часто с 2 корнями. Задние щечные зубы уменьшены в размере, как и у другого специализированного энтомофага, африканского земляного волка (Proteles cristatus). На ml метаконид в виде крупного отдельного бугорка. Палеонтологический материал крайне скуден: беззубый череп из Кангры (Kangra) в Индии (Himachal Pradesh). Размеры черепа (М. (U.) theobaldi) крупные, Пилгрим (Pilgrim, 1932) первым рассматривал его как непосредственного предка современного губача. Стратиграфическое положение находки – по-видимому, верхний сивалик (зона Boulder), т. е. верхний плиоцен. Есть ещё краниаль из пещеры Чандрапал (Chandrapalle) на юге Индии. Также из Индии есть сообщения о плейстоценовых находках, но они до сих пор не описаны.
Малайский медведь, Helarctos - укороченный лицевой отдел черепа и сильно расширенная мозговая коробка. Слуховые капсулы расширены больше, чем у остальных урсин. Зубной ряд укорочен, клыки для медведей подобных размеров гипертрофированы, метаконид на ml у заднего края протоконида. Ископаемые находки известны из позднеплейстоценовых горизонтов Вьетнама и Индонезии. Таже есть мандибулярный образец U. (Helarctos) sinomalayanus из плиоцена Китая, которые Здански и Тениус считали прямым предком нынешнего малайского медведя. Есть ещё упоминание об аптечном М2, на основе которого описывался U. praemalayanus, но в целом родственные связи малайского медведя неизвестны. В то же время, родственные связи с губачом подтверждаются морфологическим сходством и гибридизацией в зоопарковских условиях.
Надрод Ursus эволюционно наиболее прогрессивный в семействе, передние премоляры редуцированы или утрачены, протокон на Р4 смещается кзади, коренные удлинены. Нижний моляр m2 у ранних форм короче, чем ml, но впоследствии удлиняется и нередко длиннее ml.
Ursus minimus, овернский медведь – занимает базальное положение в филогенезе и, предположительно, предковый таксон для чёрных, пещерных и бурых медведей. Таксономическое положение под вопросом. Некоторые исследователи делят U. minimus на несколько таксонов: европейского раннеплиоценового U. boeckhi, европейского позднеплиоценового U. minimus, позднеплиоценового китайского U. yinanensis и раннеплиоценового американского U. abstrusus. Другие (Барышников) считают их подвидными политипичными видами U. minimus, имевшего голарктическое распределение.
В европейских образцах прослеживается эволюционные стадии смены русцинийского U. m. boeckhi (MN14-15) на ранневиллафранкского U. m. minimus (MN15-16). Что касается азиатского U. yinanensis (pанний виллафранк, MN16a) и американского U. abstrusus (нижний плиоцен, ранний бланкий, MN15), то наличествует сходство в русцинийской, азиатской и американской форме – небольшие размеры и одинарный метаконид на ml, в то время как номинативный U. minimus более крупный и молярный метаконид у него двойной.

Нижняя челюсть Ursus abstrusus
На самом деле Ursus abstrusus очень интересный мишка – по сути, небольшой примитивный чёрный медведь с длинным узким черепом, морфологически очень схожий с ранней формой U. m. boeckhi.
Череп Ursus thibetanus
Selenarctos thibetanus, гималайский медведь - сходство в дентальной морфологии предполагает, что предком являлся U. minimus, хотя конкретных доказательств пока не имеется. Плейстоценовые находки описывались под разными видовыми названиями, но дентальный статистический анализ показал, что все европейские образцы могут принадлежать к подвиду S. t. mediterraneus.
Несколько проблематична позиция Plionarctos stehlini из Мосбаха, который крупнее и, возможно, обозначает реверсивную миграцию в среднем плейстоцене. Также не вполне понятно систематическое положение индийского U. namadicus, демонстрирующего «мозаику» зубных признаков Selenarctos thibetanus, Helarctos malayanus и U. etruscus (возможно, адаптивность).
Морфометрически плейстоценовые находки Selenarctos из Закавказья (Карабах), Урала (Мохневская пещера), Китая и Японии сходны с современными формами, демонстрируя лишь относительное уменьшение хищнических зубов и удлинение задних коренных.
Euarctos americanus, барибал - первые находки относятся к раннему ирвингтонию (нижний плейстоцен). Не исключается евроазиатское происхождение, хотя достоверные находки U. americanus в Азии неизвестны.
Стратиграфически установлена тенденция к изменению размеров U. americanus во времени: сравнительно небольшие барибалы (из Port Kennedy) нижнего плейстоцена (ранний ирвингтоний) сменяются более крупными U. a. vitabilis в ср. плейстоцене (п. ирвингтоний), затем их величина в п. плейстоцене (ранчолабреаний) - U. a. amplidens (=fossilis) ещё увеличивается, а современные барибалы мельчают. К сожалению, в настоящее время наиболее диагностичен только позднеплейстоценовый материал. К слову, подвидовой состав современного U. americanus до сих пор не ревизован.
Череп и реконструкция Ursus etruscus
Ursus etruscus, этрусский медведь - лицевой отдел черепа длиннее, чем у других представителей, мозговая коробка расширена слабо, верхний хищнический зуб Р4 с крупной внутренней вершинкой, талон на М2 относительно удлинён, метаконид на ml одинарный или двойной.
Предком является U. minimus. Имеется мнение, что ранняя форма имела широкое палеарктическое распространение, в то время как на поздней стадии эволюции вида его ареал, вероятно, не выходил за пределы Европы (как и у U. minimus). Хроноподвидовые группировки: U. etruscus saintvallierensis и U. е. etruscus. U. verescagini из Таджикистана морфологически близок к ранней форме этрусского медведя, характеризуясь широкими молярами и ростральной областью. Образец из Куруксая также имеет примитивные признаки (одинарный метаконид на ml). Диагностика китайских образцов спорна, но морфологически также сходна с ранним «этруском».
P. S. По сути, филогенетически роды Melursus и Helarctos занимают близкое положение и разделяют прогрессивные характеристики – это сестринские таксоны. Палеонтологического материала почти нет, а морфо-экологическая специализация свидетельствует о формации таксонов, как минимум, уже в плиоцене. Не исключено, что и немного раньше.
Время дивергенции Selenarctos thibetanus и Euarctos americacus – ранний плиоцен. Кроме того, имеется идея о разделении чёрных медведей на Protarctos (P. boeckhi MN14, P. ruscinensis MN15, P. yinanensis MN16-17), Euarctos (E. pyrenaicus MN15, E. minimus MN16-17) и современные формы – т. е., гималайский медведь и барибал вообще исключаются из рода Ursus. Насколько в курсе, данное мнение в большей степени основано исключительно на морфологии нижних моляров, а это вызывает множество скептических вопросов. Г. Ф. Барышников объединяет Ursus minimus, Ursus etruscus, гималайского медведя и барибала как Euarctos и объективно эти формы действительно выглядят лишь как морфовариации чёрного медведя. Но обособление Protarctos как предка бурого мишки и пещерного филогенетически выглядит вроде бы оправданным.
По поводу этрусского медведя - выражены стратиграфические стадии смены хроноформ, характеризующихся уменьшением предкоренных и увеличением моляров Интересно рассмотреть образцы этрусских медведей Центральной Азии – по сути, от них и пошли адаптационные адаптации, впоследствии сформировавшие бурых и пещерных урсин.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен