Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#251 28 October 2015 17:06:52
- Алекс
- Сан Саныч - Модератор

- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические пантеры (Pantherini)
shish02 :
Сан Саныч, спасибо!
Коротко ,о чем там речь ?
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#252 28 October 2015 23:31:53
- shish02
- Гость
Re: Доисторические пантеры (Pantherini)
Алекс :
shish02 :
Сан Саныч, спасибо!
Коротко ,о чем там речь ?
Аннотация: В Европе находки гепардов (род Acinonyx), позже датируемые ранним плейстоценом, оставались ограничены некоторыми экземплярами из фауны Hundsheim (Австрия; начало среднего плейстоцена), описанными как A. intermedius THENIUS, 1954 и фрагментом плечевой кости из Mosbach Sands (Германия; начало среднего плейстоцена), который был отнесен к A. pardinensis (CROIZET et JoBERT, 1828). Здесь представлены две дополнительные находки из Мосбаха. Хотя число возможных сравнений невелико, можно предположить, что останки гепардов из обоих мест должны быть приписаны к разным таксонам. Макровидовая концепция гепардов плиоцена-плейстоцена поддерживается недавно полученными знаниями о европейских гепардах раннего плейстоцена, а также недавно описанным таксоном из Северной Африки и Северного Китая. Соответственно, гепарды, появляющиеся в Центральной Европе во время явно короткого периода начала среднего плейстоцена здесь именуются A. pardinensis (sensu lato) intermedius. Этот таксон
первоначально мог произойти в раннем плейстоцене от восточноазиатской формы A. pardinensis (s. 1.) pleistocaenicus (ZDANSKY, 1925), которая, в свою очередь, произошла от северокитайской позднеплиоценовой формы A. pardinensis (s. 1.) linxiaensis (Qm et al., 2004).
#253 29 October 2015 09:55:57
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические пантеры (Pantherini)
Андрей,спасибо ! А что за невероятные показатели длины бедренной кости 369 мм и 342 мм - это чьи ?
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#254 29 October 2015 17:27:56
- shish02
- Гость
Re: Доисторические пантеры (Pantherini)
Алекс :
Андрей,спасибо ! А что за невероятные показатели длины бедренной кости 369 мм и 342 мм - это чьи ?
Сан Саныч, с немецким лучше конечно к Игорю. Насколько я понял из текста и из подписи к планшету 342 мм - кость из Casa Frata (поздний Виллафранк, Италия), а 369 мм - из Унтермасфилда (Германия) На рис.3 (слева - Унтермасфилд, центр- Каза Фрата, справа - современный гепард) кости даны не в масштабе (у современного гепарда - 250 мм)
#255 30 October 2015 18:48:06
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Доисторические пантеры (Pantherini)
Ну, братцы, уж вы слишком большого мнения о моих лингвистических возможностях. Знания ограничены школьной программой и четырёхгодичном курсом военного училища, на котором учили вести допросы ![]()
Уважаемые коллеги, ответил в теме «Гепарды» раздела «Доисторические млекопитающие».
Саня, постараюсь обширней дополнить ответ Андрея Владиславовича…
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#256 20 December 2015 23:03:04
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Доисторические пантеры (Pantherini)
Очередное мыслеизвержение и, надеюсь, не абортированное))) по эволюционной истории пантер.
Если обратиться к наиболее свежим исследованиям, то мы получим следующие цыфэрки… Берём работу т. Кристиансена (Christiansen, 2012. Phylogeny of the sabertoothed felids (Carnivora: Felidae: Machairodontinae)) по филогении махайродонтов и получаем расхождение между ними и коническозубыми кошками примерно 17 млн. Это с учётом, что саблезубые кошаки группы миомахайродов, которые, по сути, настоящими махайродами ещё не являются, фиксируются только в среднем миоцене, примерно 12 млн лет назад (поздний астараций Турции (MN 8), Machairodus robinsoni).
Отложим статью достойного потомка викингов и обратимся к работе китайского коллеги Ценга и его бригады (Tseng et al., 2013. Himalayan fossils of the oldest known pantherine establish ancient origin of big cats), опубликованную с описанием Panthera blytheae. Здесь темпоральные границы более «размыты» - средние значения 16,4 млн с формированием базального рода 10,7 млн, однако допустимые значения появления фелид пантеринной группы аж 27,6 млн (!!!).
Обратимся к палеонтологическим данным. Pseudaelurus, как уже истинная кошка, известна из биозоны MN3 (20,5-18,0 млн). Ряд форм описан из раннего миоцена Евразии, Африки и Северной Америки. Не вдаваясь в морфологические нюансы образцов псевдайлуроидных кошек, достойные отдельной дискуссии, стоит отметить, что истинных фелин определяют уже в раннемиоценовых отложениях. И если Diamantofelis и Namafelis ещё близки к псевдайлурусам, то Asilifelis (17,5 млн лет назад, Намибия) занимает промежуточное положение между таковыми и кошками фелинной градации. А в конце среднего миоцена (12 млн) уже появляются явные среднеразмерные фелины Styriofelis и Pristifelis. Североамериканские таксоны типа Pratifelis и Hyperailurictis во внимание не берутся.
Номенклатурное обозначение рода Panthera и его выделение в отдельное подсемейство в начале XX века было весьма дискуссионным, но в конце концов г. Покок позиционировал его отдельным таксоном (Pocock, 1917. The classification of the existing Felidae) на основе морфологии подъязычного аппарата и ряда других характеристик – правда, не очень чётко обозначенных. Был ещё такой парняга по фамилии Гершковиц, который с ним спорил. Стоит отдать должное, чувак был из «полевиков», но изучал ВСЁ и лез ВЕЗДЕ – от сумчатых до китообразных и всюду считал должным доминировать. Но в итоге справедливость восторжествовала.
Логически рассуждая, принимая во внимание структуру гиоидного аппарата, пантерины не могли появиться позже фелинных кошек. Однако с миоценовыми фелинами Азии не всё так просто. Среднемиоценовый Pseudaelurus chinjiensis вроде определён в Vishnufelis, но насколько сейчас данный таксон валиден, не в курсе. Явные псевдайлуроиды фиксируются в китайских местонахождениях: Pseudaelurus cuspidatus раннего миоцена и P. guangheensis и P. lorteti среднего. Вот где-то здесь, наверное, и следует искать предковые формы архаичных фелид, локально обособленные популяции которых сперва соматически, затем феноадаптивно а, впоследствии, путём геновариаций и сформировали морфотип пантерин.
Посему idea fix – гиоидный аппарат дымчатого леопарда. Знаком с его структурой у пантер (ягуара, льва, тигра) и фелин (гепард, домашняя кошка). Анатомическое описание этих сегментов у Neofelis нигде не могу найти…
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#257 21 December 2015 00:37:55
- vas
- Любознательный
- Зарегистрирован: 29 May 2014
- Сообщений: 120
Re: Доисторические пантеры (Pantherini)
Сorvin
Посему idea fix – гиоидный аппарат дымчатого леопарда. Знаком с его структурой у пантер (ягуара, льва, тигра) и фелин (гепард, домашняя кошка). Анатомическое описание этих сегментов у [i :Neofelis[/i] нигде не могу найти…
Clouded leopards have an ossified hyoid that allows them to purr but not roar. They also can make a very distinctive chuffing sound as well as other vocalizations.
http://www.cloudedleopard.org/documents … manual.pdf
Неактивен
#258 21 December 2015 12:18:44
- shish02
- Гость
Re: Доисторические пантеры (Pantherini)
При написании истории рода Panthera или подсемейства Pantherinae ориентироваться на гиоидный аппарат (и даже уделять ему сколь-нибудь большое значение), мне кажется, не стоит. Подъязычная кость в ископаемом состоянии практически никогда не сохраняется (не говоря уже о всем гиоидном аппарате). Известна ли подъязычная кость у Panthera blytheae? Нет. Равно как неизвестна она и у африканских леоподобных кошек позднего плиоцена. Тем более, как показывает история с дымчатым леопардом, хищник может не иметь способности рычать, но при этом относится к Pantherinae на основе других морфологических характеристик.
Могут ли здесь помочь генетики, например, рассчитав примерное время формирования данного аппарата у пантер? Не уверен. Когда речь идет о позднем миоцене (и уж тем более среднем миоцене) расчеты генетиков начинают носить все более спекулятивный характер (грубо говоря, можем ли мы поручиться за то, что скорость эволюционных изменений у фелид миоцена, в период интенсивного образования даже не новых видов, а новых родов, была точно такой же как и у современных кошек?). Печальный факт состоит в том, что между появлением первых среднеразмерных фелид, которые насколько я понимаю, не относятся даже к Pantherinae, и находками первых пантерин (Panthera blytheae) мы имеем ничем не заполненный разрыв почти в 6 млн. лет.
Определить какой вид псевдайлуроидных кошек дал начало линии, ведущей к подсемейству Pantherinae мне не представляется возможным. Даже самые прогрессивные представители рода Pseudaelurus могли с одинаковым успехом дать начало как линии, ведущей к роду Panthera, так и к линии, ведущей к любому другому роду фелид. Между псевдайлуринами и современными кошками существовали промежуточные формы, о которых мы почти ничего не знаем ![]()
#259 21 December 2015 18:09:58
- shish02
- Гость
Re: Доисторические пантеры (Pantherini)
В работе Христиансена "SPECIES DISTINCTION AND EVOLUTIONARY DIFFERENCES IN THE CLOUDED LEOPARD (NEOFELIS NEBULOSA) AND DIARD’S CLOUDED LEOPARD (NEOFELIS DIARDI)" обнаружил интересный момент. Если совсем кратко, то в работе весьма убедительно обосновывается, что с т.з. краниальной морфологии и краниометрии различия между Neofelis diardi (островные популяции дымчатого леопарда) и Neofelis nebulosa (материковые популяции дымчатого леопарда) соответствуют не подвидовому, а межвидовому уровню (в качестве контрольной группы выступали леопарды, ягуары и тигры). Это-то все хорошо и замечательно, но вот что автор пишет далее: "Исследование молекулярных данных наводит на мысль о репродуктивной изоляции N. diardi от материковых дымчатых леопардов приблизительно 1,4-2,9 млн. лет назад, что сопоставимо с видовой изоляцией у других пантерин (Buckley-Beason et al. 2006; Wilting et al. 2007).
В раннем плейстоцене (до 0,8 млн. лет назад) Ява, по-видимому, действительно была изолирована от континента (van den Bergh GD, de Vos J, Sondaar PY (2001) The Late Quaternary palaeogeography of mammal evolution in the Indonesian Archipelago. Palaeogeogr Palaeoclimatol Palaeoecol 171 р.390). Тут надо оговориться, что данный вывод сделан скорее на основе косвенных признаков (наличие мелких элефантид, мелких стегодонтов и гигантских черепах - "эта ассоциация предполагает островное положение Явы в начале Четвертичного периода"). Но была ли изолирована Суматра? Кажется, нет. Как пишет Тоугард (Tougard) в работе "Biogeography and migration routes of large mammal faunas in South-East Asia during the Late Middle Pleistocene: focus on the fossil and extant faunas from Thailand" до 0,8 млн. лет назад колебания моря были умеренные, со средним значением 70 м ниже уровня моря в наши дни (максимум - 100 м). В настоящее время, для соединения Суматры, Явы и Борнео с материком достаточно падения уровня моря на 50 м (см. Тоугарда). Что-то не сходится с островной изоляцией или точнее с ее хронологическими рамками.
Отредактировано shish02 (21 December 2015 18:40:20)
#260 22 December 2015 22:33:59
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Доисторические пантеры (Pantherini)
Vas и Андрей, спасибо! Я об этом и подозревал.
Андрей Владиславович, как мне кажется, морфологию эпигиоида при дифференциации пантер всё же не стоит совсем сбрасывать со счетов. У фелин он костистый, у снежного барса – промежуточная морфология, у тигров, львов, ягуаров и леопардов это упругая связка. Кроме того, топология элементов подъязычного аппарата у последних изменяется онтогенетически, с возрастом. Связано ли это с вокализационными проявлениями общих бихевиористических аспектов представителей группы (обозначение территории, течка, угроза конкурентам и т. д.) или проявление некоего анатомического атавизма? У тигров и львов фактическое участие акустической демонстрации низкочастотных звуков в поведении несомненно. С другой стороны, эволюция слепа и её многие соматические огрехи мы испытываем на собственной шкуре – аппендикс, зубы «мудрости», расположение гланд в глотке и т. п. Если был бы Бог, то он как биоконструктор модернизировал бы нас более прогрессивно))).
То, что структура подъязычного аппарата вариативна и имеет тенденцию к синостозии (сращению) наблюдается как у различных представителей пантерин (у ягуаров, львов и леопардов он из весьма эластичных связок, а у тигров связь между подъязычным аппаратом и гортанью более синовиальна), так и в онтогенезе – у взрослых особей пантер вся эта система занимает более каудальную позицию при более удлинённом зеве, чем у ювенильных, у которых общая топография гиоидного аппарата сходна с фелинными кошками: «базигиоид, тиреогиоид и прилагаемая гортань изменяют своё положение от рострального до каудального в ходе последующего развития у пяти видов из семейства кошачьих вместе со связками эпигиоида - то есть, у льва, леопарда, ягуара, тигра и снежного барса» (Weissengruber et al., 2002, "Hyoid apparatus and pharynx in the lion (Panthera leo), jaguar (Panthera onca), tiger (Panthera tigris), cheetah (Acinonyx jubatus) and domestic cat (Felis silvestris, f. catus)".
Фактически, морфология подъязычного аппарата, глотки и их топографических позиций свидетельствует о следующем:
1. Виды, объединённые под родовым именованием Panthera – монофилетичны.
2. Ягуар, лев и леопард морфофизиологически имеют некоторую дифференциацию от тигра.
3. Оссификация элементов гиоидного аппарата изначально заложена в морфофизиологии фелид – очевидно, ещё на уровне представителей прогрессивных псевдайлурин. Онтогенетически это демонстрируется у вскрытых ювенильных и половозрелых ягуаров. Филогенетически – у базальных пантерин типа Neofelis и признак, имеющий тенденцию к остеогенезу у снежного барса Panthera uncia.
С этим вроде чуть разобрались. Вообще, очень даже желается суммировать отличительные признаки в диагностическую матрицу. Пусть она не является непреложным аргументом, но в сравнительном анализе полезна.
Андрей Владиславович, клёво, что мы солидарны в вопросах генетических изысканий. На нынешнем уровне молекулярной генетики в вопросах изучения ископаемых образцов более-менее адекватные выводы могут быть только в хронодиапазоне голоцена-позднего плейстоцена. Какой там на хрен плиоцен и, тем более, миоцен? Андрюш, да эти предположения ещё более фантастичны, чем тот проект, который мы замыслили на методологической основе в теории – диалектических, на практике – морфофизиологических методах. Евгений Гасанов (Женёк, извини, но отчества твоего не знаю) уже формулировал методы и результаты современных генетических изысканий. При всём прогрессе нынешних результатов - в палеонтологии они в объективном плане мизерны и фантастичны.
Проблематичны и вопросы морфологии. В середине прошлого века Хеммер предполагал, что Pardofelis и Neofelis относятся к группе пантерин. «Облачный леопёрд» сейчас в базальной позиции, мраморная кошка генетиками обособлена. Надеюсь, здесь в какой-то степени всё же поможет матрица.
Андрей, другой вопрос, который ты предвосхитил – темпы эволюционного развития и низкий тебе за это поклон, дуем в одну дудку. Значицца, дело пойдёт, потому что я тоже в теорию видообразовательных процессов пантер эту константу вменял. Я абсолютно и непреложно уверен в том, что дивергенция пантер и фелин происходила одновременно. Примерно на рубеже раннего-среднего миоцена, около 18 млн лет назад. Первые фелины уже достаточно распространены в евроазиатском регионе. Следовательно, ареал протопантерин был весьма ограничен. Очевидно, тропиками южной и юго-восточной Азии. Вообще, выглядит так, что архаичные формы пантерин «варились» в биогеографически обособленном котле (возможно, изначально даже ограниченном локально), а потом «выстрелили». По-видимому, в процессе адаптации освоив экологическую нишу лесостепных биотопов (будучи сперва трофически если не угнетённой, то весьма обособленной группой), а с расширением данной зоны на миоцен-палеоценовой границе начав интенсивную радиацию.
Андрюш, поэтому я и включил в эволютивные факторы биогеографический контекст. Скорость эволюции пантер зависела в первую очередь от абиотических факторов. И её макромасштабность в плане видо- и родообразования напрямую была связана с эволюцией экосистем. Каким образом? Пока не знаю, представляя лишь "генерально", а подробнее буду копаться. Подспудно отринув субъективность )))). Ведь самому интересно…
В отношении дымлеопёрда… Статья пользительная, бесспорно. Только зачем струнить растяжку в духовке, если гроздь Ф-1 можно насторожить уже на входе в кухню? Иными словами – зачем постулировать сенсацию на пустом месте? Где серьёзный анализ обособления островных популяций? Мля, об этом ышшо дедушка Дарвин писЫвал. А здесь попахивает мазаковщиной, когда разделяли тигров на два вида… Но интересные моменты в работе имеются, которые следует взять на вооружение, здесь товарищу Перу агромадное спасибочко.
Географией Сунда как-то интересовался, хотя и давненько, когда в тигров и леопардов Индокитая «втыкался». Регрессия там, особенно в конце среднего-позднем плейстоцене довольно динамичной была но, по-моему, собственно Сунд как тихоокеанский сухопутный псевдорегион был только в среднем плейстоцене (первый грандиозный ледниковый максимум примерно 660-650 млн лет назад), захватывая Борнео. А связь полуострова Малакка с Явой и Суматрой ещё в плиоцене фиксируется.
В общем, надо лазать по букварям.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#261 16 January 2016 15:00:25
- Crazy Zoologist
- Гость
Re: Доисторические пантеры (Pantherini)
Что за странные данные?
The 235 kg average has no basis, as Guate confirmed. The specimens we have average around 300 kg in my report, albeit there is a significant difference between the 470 kg specimen (even though there are 7 fossils, this one is an outlier) and the other two males estimated in 270-320 kg range specimens. Don't get too confused by the difference the numbers Guate and I give, it's due to myself using some extra measurements of the fossils I took myself and my own equations from databases in my area. Point is that 235 kg is much too low no matter which way you look at it.
Here's a summary of all mass estimates from my database. Greatest mass in parenthesis.
At the moment, off length dimensions only, the ranks are as follows:
1. Ngandong tiger - Femoral length (409 kg)
2. Cromerian lion - Ulna Length (400 kg)
3. American lion - Femoral Length (350 kg)
3. Cave lion was the same size as the modern lion in this measurement
4. Smilodon populator was the same size as the modern lion and tiger in this measurement
Off width dimensions only:
1. Ngandong tiger - Femoral DAW (531 kg)
2. American lion - Femoral DAW (500 kg)
3. Smilodon populator - Humeral DAP (497 kg)
4. Cromerian lion - no width dimensions available for largest specimen, although femur data shows a species with slightly thinner bones than the modern lion
5. Cave lion - Slightly larger than modern lion
Taking into all measurements:
1. Ngandong tiger (470 kg)
2. Smilodon populator (440 kg)
3. American lion (415 kg)
4. Cromerian lion (400 kg)
5. Cave lion (272 kg)
Previously, I was under the assumption that P. atrox was a bit smaller than P. fossilis. But after comparing diameters, the largest of P, atrox (450 mm of massive DAW=120 mm), edges P. fossilis.
In terms of length measurements, all species are assumed to be as dense as tigers. In terms of width, only the Ngandong tiger was given an only tiger database for estimation, as tiger's are slender boned for their density. Smilodon was shared with bears. The "lions" were based on a mix of tigers and modern lions.
Cromerian lion что за лев? Кто-то из плейстоценовых.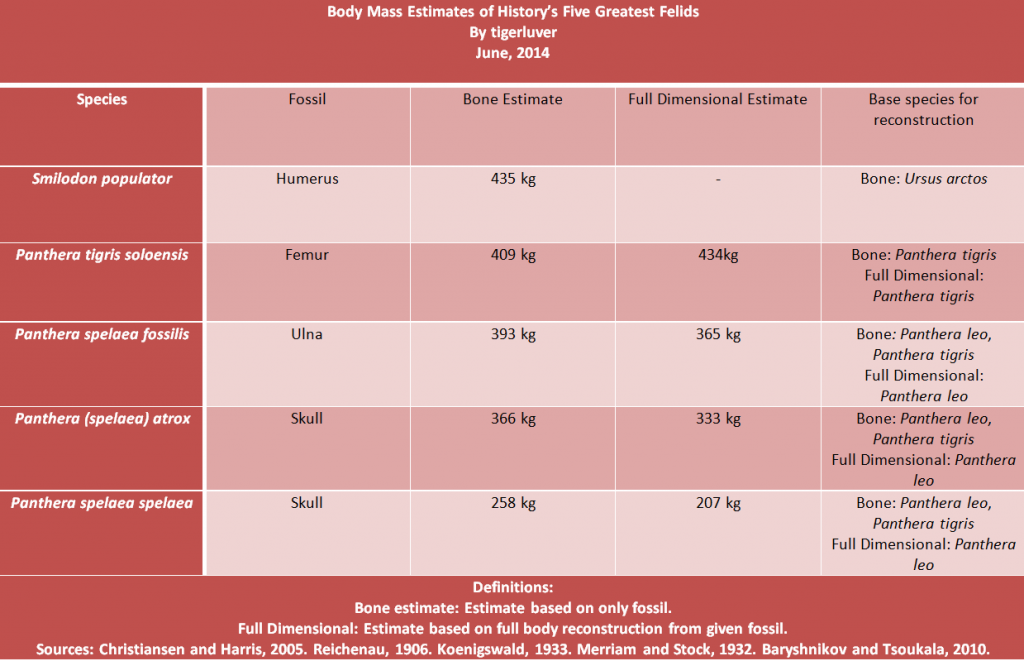

Отредактировано Амфицион (16 January 2016 15:02:40)
#262 16 January 2016 15:27:15
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические пантеры (Pantherini)
Амфицион :
Что за странные данные?
The 235 kg average has no basis, as Guate confirmed. The specimens we have average around 300 kg in my report, albeit there is a significant difference between the 470 kg specimen (even though there are 7 fossils, this one is an outlier) and the other two males estimated in 270-320 kg range specimens. Don't get too confused by the difference the numbers Guate and I give, it's due to myself using some extra measurements of the fossils I took myself and my own equations from databases in my area. Point is that 235 kg is much too low no matter which way you look at it.
Here's a summary of all mass estimates from my database. Greatest mass in parenthesis.
At the moment, off length dimensions only, the ranks are as follows:
1. Ngandong tiger - Femoral length (409 kg)
2. Cromerian lion - Ulna Length (400 kg)
3. American lion - Femoral Length (350 kg)
3. Cave lion was the same size as the modern lion in this measurement
4. Smilodon populator was the same size as the modern lion and tiger in this measurement
Off width dimensions only:
1. Ngandong tiger - Femoral DAW (531 kg)
2. American lion - Femoral DAW (500 kg)
3. Smilodon populator - Humeral DAP (497 kg)
4. Cromerian lion - no width dimensions available for largest specimen, although femur data shows a species with slightly thinner bones than the modern lion
5. Cave lion - Slightly larger than modern lion
Taking into all measurements:
1. Ngandong tiger (470 kg)
2. Smilodon populator (440 kg)
3. American lion (415 kg)
4. Cromerian lion (400 kg)
5. Cave lion (272 kg)
Previously, I was under the assumption that P. atrox was a bit smaller than P. fossilis. But after comparing diameters, the largest of P, atrox (450 mm of massive DAW=120 mm), edges P. fossilis.
In terms of length measurements, all species are assumed to be as dense as tigers. In terms of width, only the Ngandong tiger was given an only tiger database for estimation, as tiger's are slender boned for their density. Smilodon was shared with bears. The "lions" were based on a mix of tigers and modern lions.Cromerian lion что за лев? Кто-то из плейстоценовых.
http://i293.photobucket.com/albums/mm56 … g~original
https://lh3.googleusercontent.com/_sBJX … 72-h555-no
http://forum.zoologist.ru/viewtopic.php … 2&p=19 сообщение 465 таблица .
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#263 16 January 2016 15:35:50
- Crazy Zoologist
- Гость
Re: Доисторические пантеры (Pantherini)
Ну это понятно, что это бред. Христиансен об этом и пишет.
А кто такой Cromerian lion?
#264 16 January 2016 16:54:26
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические пантеры (Pantherini)
Амфицион :
Ну это понятно, что это бред. Христиансен об этом и пишет.
А кто такой Cromerian lion?
наверное фоссилис.
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#265 16 January 2016 17:01:32
- Crazy Zoologist
- Гость
Re: Доисторические пантеры (Pantherini)
А к какому подвиду отнесены плейстоценовые львы Африки? Почему так мало останков? Че-то мне кажется, что львы современного типа образовались уже в голоцене. Недаром лев считается более прогрессивным видом, чем тигр, который в позднем плейстоцене был таким же как в голоцене. И вообще он более кошачий.
#266 16 January 2016 21:10:00
- shish02
- Гость
Re: Доисторические пантеры (Pantherini)
Амфицион :
Ну это понятно, что это бред. Христиансен об этом и пишет.
А кто такой Cromerian lion?
Кромерский комплекс (по североевропейской стратиграфической шкале) датируется примерно 850-600 тыс. лет. Ну, и соответственно, кромерский лев - это тот же фоссилис
#267 01 February 2016 07:12:19
- Юра
- Без пяти минут зоолог
- Зарегистрирован: 31 July 2008
- Сообщений: 10735
Re: Доисторические пантеры (Pantherini)
Череп Panthera blytheae в ЗD:
http://phenome10k.org/panthera-blytheae/
Неактивен
#268 01 February 2016 12:43:56
- Crazy Zoologist
- Гость
Re: Доисторические пантеры (Pantherini)
Юра :
Череп Panthera blytheae в ЗD:
http://phenome10k.org/panthera-blytheae/
А где он найден и каким возрастом датирован? Промеры черепа есть?
#269 01 February 2016 14:07:16
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Доисторические пантеры (Pantherini)
Амфицион :
Юра :
Череп Panthera blytheae в ЗD:
http://phenome10k.org/panthera-blytheae/А где он найден и каким возрастом датирован? Промеры черепа есть?
http://rspb.royalsocietypublishing.org/ … 4/20132686 Артем ,ленивый ты стал ,все есть в википедии ![]()
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#270 01 February 2016 15:47:28
- Crazy Zoologist
- Гость
Re: Доисторические пантеры (Pantherini)
А, так эта та самая древняя якобы пантерка, родственник ирбиса? Читал про нее, но латынь забыл.
#271 10 March 2016 16:47:08
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Доисторические пантеры (Pantherini)
Некоторые соображения по филогении ягуаровых кошек, извините за «топорность» исполнения:
Panthera sр. – гипотетический азиатский трихотомичный предок ягуаров, леопардов и львов.
Panthera georgică – грузинская пантера (правая ветвь нижней челюсти с зубным рядом р3-m1, no. 2027, Национальный музей Грузии, Тбилиси), морфологически схожая с европейскими ягуароидами группы toscana. Обозначает как одну из самых ранних находок ягуароподобных пантер, так и географически подтверждает миграцию таксона из Азии в Европу в первой трети раннего плейстоцена.
Panthera toscana – первый ягуароморф. Некоторыми исследователями считается подвидом гомбасцогской пантеры, однако сравнительный морфологический и морфометрический анализ позволяет сделать вывод, что европейские палеоягуары всё же разделяются на группы позднего виллафранка (P. toscana) и ягуароподобных пантер конца раннего-среднего плейстоцена (P. gombaszoegensis).
Panthera gombaszoegensis ssр. – переходная (toscana-gombaszoegensis) хроноформа гомбасцогской пантеры из эпивиллафранка Европы, демонстрирующая отличительные дентомандибулярные особенности. Некоторыми (Mol et al., 2011; Marciszak, 2013) таковые рассматриваются в качестве индивидуальной изменчивости или полового диморфизма, однако эти признаки (высота нижнечелюстной ветви, длина заклыковой диастемы, ширина черепа в лобной области, морфология и размеры р3 и р4 и т.д.) являются общими для образцов на географически довольно обширном районе – от Нидерландов (Maasvlakte) до Италии (Vallonet) и длительными стратиграфически (1,1-0,8 млн), что свидетельствует о прогрессивных изменениях в группе, которые достойны обозначения хотя бы на уровне хроноподвида. Типовым материалом могут послужить образцы из германского местонахождения Унтермассфельд.
Panthera gombaszoegensis gombaszoegensis – архетипичная форма среднего плейстоцена.
Panthera (gombaszoegensis) ssр. (transcaucasiсa ?) – транскавказский морфотип, сходный с ранней эпивиллафранкской P. gombaszoegensis. «Узловой» таксон, наметивший прохорез ягуароидов в сторону Нового Света в конце раннего плейстоцена. География находок подтверждает масштабность экспансии; более поздний материал из Кударо и Саудовской Аравии – или реликты, или же повторная миграционно-аппендиксная волна.
Panthera (оnca) augusta – североамериканский плейстоценовый ягуар. Таксономическая позиция под вопросом: с одной стороны, отличия от современных подвидов не очень значительны; с другой – посткраниальные пропорции, да и индексальные значения черепа свидетельствуют о том, что экстерьерно он выглядел более «леоподобно», чем P. оnca. Некоторые дентальные морфологические отличия также присутствуют (в диапазоне вариаций современных форм), однако обстоятельный анализ с обширной выборкой и учётом как хронологии, так и географических рас никем не проводился.
Panthera (оnca) augusta ssр. – выделен в качестве хроноподвида типовой формы ввиду явного уменьшения общего размера и пропорции конечностей ягуаров висконсина в сравнении с иллинойскими образцами. Стоит учесть, что хронологическая разница с вышеописанной формой почти в полмульёна лет.
Panthera (оnca) mesembrina – также проблематичный таксон с точки зрения систематики. Среднеплейстоценовые остатки (энсенадий) гомологичны P. (оnca) augusta Северной Америки, позднеплейстоценовые (луханий) – современному P. оnca. Возможно, выделявший последнюю филогенетическую ветку как P. o. palustris Сеймюр (Seymour, 1989) таки прав, но здесь следует учитывать невообразимое смешение хроноподвидов, географических популяций, возрастных, гендерных и индивидуальных различий… Хотя бы, к примеру, потому, что эквадорские ископаемые ягуары отличаются от аргентино-базильских (Cordy, 1982) А вообще, пусть они разбираются со своими кошаками сами.
Panthera оnca – без комментариев. Единственно – филогения современных подвидов векторизована в соответствии с последними изысканиями, основанными на данных генетических анализов (Eizirik et al., 2001). А вот подробных, обширных, обстоятельных и фундаментальных морфоисследований также до сих пор не имеется.
P.S. Большое спасибо Андрею Владиславовичу за косвенную помощь.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#272 10 March 2016 19:15:23
- shish02
- Гость
Re: Доисторические пантеры (Pantherini)
Игорь, спасибо за труд. Все здорово. Немного добавлю от себя. Panthera (оnca) augusta представляется вполне валидным таксоном. Собственно говоря плотно им занимались только Куртен и Сеймур (нет, есть и другие исследователи, но их работы обычно не идут дальше находок в одном конкретном месте). Так вот эти два матерых человечища данный таксон вполне себе признавали. Им, думаю, в данном случае и следует доверять. Ни о каких расах при таком количестве самых разномастных находок, конечно же, и речи нет. Опять же, в принципе, можно нарисовать-то по принципу юг-север или восток-запад, но цена этим выделенным расам при такой выборке и индивидуальной изменчивости будет грош в базарный день.
Насколько я уловил последний тренд, сейчас принято всех южноамериканских плейстоценовых ягуаров именовать Panthera (оnca) mesembrina. Конечно, это все не строго, некоторые исследователи вообще с подвидами не заморачиваются и пишут просто - Panthera оnca, даже если речь о среднем плейстоцене. У Сеймура P. o. palustris и Panthera (оnca) mesembrina рассматриваются как синонимы. На том и сойдемся. Подвид (на мой сугубо дилетантский взгляд вполне валидный) интересен еще и тем, что на протяжении плейстоцена не уменьшался в размерах, как Panthera (оnca) augusta, и вплоть до вымирания сохранял размеры, соответствующие крупным ирвингтонским ягуарам Сев.Америки. Кроме того, от современного ягуара отличался и с т.з. палеоэкологии.
p/s. Игорь, а чего там на счет работы Cordy, 1982? Я чего-то ее не припомню. Будь добр, скинь по почте если есть.
#273 11 March 2016 01:12:27
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5943
Re: Доисторические пантеры (Pantherini)
Андрей Владиславович, я Корди с Хоффстеттером перепутал, пардонь; высылаю обе работы, а заодно классическое описание цимбальского образца Верещагина.
Андрюш, валидность augusta и mesembrina (понятно, что они принадлежат одной филетической "ветке") меня интересует только со следующей позиции: отдельного видового статуса ягуроморфных пантерин или же подвидовой ранг? Меня устраивает и более прозрачно именно (оnca) augusta/mesembrina. Понятно, что я не Куртен и не Сеймюр, но в такой интерпретации, на мой взгляд, филогенетическая позиция плейстоценовых ранних ягуаров лучше очерчена... Ещё не совсем ягуары, но уже и не гомбасцогские пантеры)))
А mesembrina по-моему, всё же уменьшалась. Сейчас аргументировать не буду, лучше Кабреру почитать, которого нам с тобой Велизар Семионовски отфоткал (Vas). Давай, я тоже инфу поштудирую, а там пообщаемся. Кстати, ещё имеется изумительная классическая, хоть и архаичная, но фундаментальная работа Покока по ягуарам. У меня где-то в "загашниках" есть, стопудово. Также вышлю, как найду.
Ну, а так... Североамериканский Panthera (оnca) augusta издох, как это не прискорбно. А вот южноамериканский P. (оnca) mesembrina в лухании трансформировался в современного ягуара. Дальнейшая радиация в Северную Америку была реверсивной; причём, наверное, несколькими миграционными волнами. Учитывая матримональную бихевиористику пантерин, это вполне нормально.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#274 23 March 2016 15:09:20
- shish02
- Гость
Re: Доисторические пантеры (Pantherini)
Ниже представлен черновой вариант текста, где рассматривается палеоэкология плейстоценовых ягуаров. В скобках идет ссылка на источник (первая цифра - номер источника, вторая - номер страницы). Приветствуются аргументированные замечания и дополнения.
Палеоэкология плейстоценовых ягуаров
Современные ягуары имеют преимущественно тропическое распространение, стараясь выбирать густые заросли около воды. Самая крупная на сегодняшний день популяция обитает в дождевых лесах Амазонии. Дождевых лесов придерживаются и ягуары, обитающие на юго-востоке Бразилии, в районе атлантического побережья (30). Panthera onca отлично себя чувствует в болотах Пантанала. Вместе с тем, ягуары демонстрируют достаточно высокую пластичность, позволяющую им заселять и другие биотопы. Хотя современные ягуары редко поднимаются в горы на высоту более 1000 м, были зафиксированы случаи их обнаружения и на высотах в 2000 м в Венесуэле, 2100 м в Перу и 2700 м в Боливии (10,2). Предпочитая густую растительность и околоводную среду, ягуары, тем не менее, способны осваивать и засушливые районы. Даже в историческое время (до 18 века) Panthera onca был достаточно широко распространен по всему югу США, от Флориды до Калифорнии (31,467). И хотя засушливые районы Аризоны и Нью-Мексико, очевидно, нельзя отнести к предпочитаемой среде обитания ягуара, он на протяжении тысяч лет существовал и там (32). Вместе с тем, даже в не самых благоприятных для себя ландшафтах ягуар, по-видимому, стремился выбирать места возле водоемов. Так в Патагонской степи находки ягуаров имеют привязку к основными рекам (34,39). Ягуары позднего плейстоцена, найденные в пещере Gruta do Urso (Токантис, Бразилия) также придерживались околоводной среды обитания. Об этом говорят останки сопутствующей фауны (35). В прошлом ареал ягуара был гораздо шире, простираясь с севера на юг от штата Вашингтон в США до южной Патагонии в Чили. С востока на запад ареал ягуара простирался от океана до океана. Естественно, что при таком широком ареале среда обитания плейстоценовых ягуаров была более разнообразной, чем у их современных потомков. В прошлом они встречались в местах, где сейчас уже не встречаются. Например, в Mullen II (Небраска, США) останки сопутствующей ягуару фауны (Symbos, Cervalces), по мнению исследователей, могут указывать на наличие в северной части Небраски в позднем Ирвингтоне еловой тайги (19,96). Следовательно, на рубеже среднего и позднего плейстоцена ягуар обитал в среде, в которой в настоящее время обитают северные популяции пумы. По-видимому, в прошлом многие популяции ягуаров длительное время придерживались экотонов, своеобразных зон перехода (от леса к степи и т.д.), которые, обычно, характеризуются большим биологическим разнообразием. Данное обстоятельство обеспечивало ягуару пластичность и способствовало его выживанию в период массового вымирания мегафауны в конце плейстоцена.
Исследователи уже давно замечали, что современный ягуар (Panthera onca), по сравнению с другими крупными кошками, очень часто охотится на добычу, которая существенно меньше его самого. Обладая очень мощными клыками и робустным телосложением, ягуар мог бы регулярно охотиться на более крупную добычу, чего он, по-видимому, не делает. Средняя масса предпочитаемых жертв у ягуара составляет 32 (+/- 13) кг. Отношение средней массы хищника к массе жертвы у ягуара (1:0,53) существенно отличается от такового у других крупных одиночных фелид.
Рисунок, демонстрирующий отношение массы тела хищника к массе тела жертв. Источник: Hayward M.W. et al. (2016) Prey Preferences of the Jaguar Panthera onca Reflect the Post-Pleistocene Demise of Large Prey // Frontiers in Ecology and Evolution. Vol.3, Article 148. p. 13.
Последние исследования показывают, что ягуары предпочитают капибар и муравьедов. Они часто употребляют в пищу черепах и кайманов. При этом избегают охотиться на приматов, тапиров, опоссумов, других хищников (1,8). Хэйвард и его коллеги показали, что при отборе жертв современный ягуар скорее руководствуется социо-экологическими и поведенческими особенностями добычи, чем ее морфологическими характеристиками (размер, масса). По их мнению, в случае с ягуаром мы можем наблюдать «ограниченный эффект влияния морфологии жертвы на отбор добычи» (1,1). Эта особенность в сочетании с «отсутствием предпочитаемой охоты на травоядных Cetartiodactyla привела, по мнению исследователей, к редукции массы у плейстоценовых ягуаров. Ученые пришли к выводу, что вымирание в конце плейстоцена крупной потенциальной добычи наложило существенный отпечаток на хищническую экологию ягуара, и эффект от последствий массового вымирания мегафауны все еще сказывается на хищническом поведении Panthera onca. Безусловно, ягуар может успешно охотиться на довольно крупную добычу вроде оленя Марша, тапира или крупных домашних животных вроде лошадей и коров. Но эти крупные животные не являются его основной предпочитаемой добычей. Вероятно, именно переход на мелкую и весьма разнообразную добычу (в рацион современного ягуара входит, по меньшей мере, 85 видов животных, большая часть из которых существенно меньше его самого) и способствовал выживанию ягуара. В плейстоцене в Новом Свете существовало более 50 дополнительных видов травоядных животных, чья масса превышала 40 кг, и которые могли служить потенциальной добычей ягуара. Голоценовая фауна Южной и Центральной Америки характеризуется крайне низким разнообразием крупных травоядных. Это отчасти объясняет, почему современные ягуары предпочитают не ориентироваться преимущественно на размер жертвы.
Тапиры сумели пережить плейстоценовое вымирание, но их плотность, по-видимому, оказалась недостаточной для того, чтобы ягуары стали рассматривать их как основную предпочитаемую добычу. Этому, вероятно, мешала и конкуренция со стороны людей и то, что взрослые тапиры (150-300 кг) в целом выходят за размерные рамки предпочитаемой добычи. Ситуация с пекари несколько иная. По размерам они отлично укладываются в рамки предпочитаемой добычи, но таковой не являются. Исследователи предполагают, что дело в том, что ягуар стремится избежать ран, которые может нанести ему взрослый пекари. Сохранение здоровья особенно важно для одиночного хищника или самки с детенышами. Другое объяснение этого несоответствия заключается в возможном прохождении пекари через «бутылочное горло» (известно, что некоторые виды пекари не пережили массового вымирания в конце плейстоцена) и их численность резко сократилась на рубеже плейстоцена и голоцена (1,15). Позже, рассматривая североамериканских плейстоценовых ягуаров, мы покажем, почему, на наш взгляд, второе объяснение выглядит более убедительным. Высокий уровень охоты на кайманов (62 % смертей кайманов приходится на ягуаров) еще не означает, что они являются его предпочитаемой добычей. В некоторых случаях рептилии могут составлять до половины диеты ягуара. Но проведенный анализ 19 исследований позволил Хэйварду и его коллегам прийти к выводу, что в реальности это отношение около 21 % (1,15). Что касается крупного рогатого скота, то исследования показывают, что ягуар предпочитает нападать на молодых животных (до 1 года), весящих менее 120 кг. Нападений же на взрослых животных он избегает.
Мы не ставим целью подробное рассмотрение вопросов пищевой экологии и охотничьего поведения современного ягуара. Литература по данным вопросам весьма велика (2) и охватывает самые разнообразные аспекты пищевой экологии и охотничьего поведения Panthera onca. Анализ научных работ однозначно говорит в пользу изумительной пластичности данного хищника. Но если вопросы пищевой экологии современного ягуара изучены достаточно хорошо, то о палеоэкологии плейстоценовых ягуаров мы знаем гораздо меньше. Хэйвард и его коллеги подчеркивают, что «не ясно, предпочитали ли ягуары охотиться на относительно мелкую добычу в плейстоцене, или же просто использовали переход на более мелкую добычу в голоцене как стратегию выживания в конце плейстоценового вымирания мегафауны» (1,12). Первый вариант исследователям отвергается. При высоком разнообразии крупных травоядных, сравнимым с разнообразием в африканской саванне, «кажется невероятным, что ягуары ограничивались лишь мелкой добычей» (1,12). К охоте на более крупную добычу в плейстоцене, по их мнению, подталкивал и более крупный размер плейстоценовых ягуаров. Переход на более мелкую добычу, как стратегия выживания, сопровождался, по мнению Хэйварда, уменьшением размеров самих ягуаров (1,14). Такое сочетание, по мнению исследователей, не является чем-то уникальным и наблюдалось у голоценовых койотов. Переход на мелкую добычу, по мнению Хэйварда и его коллег, во многом был облегчен тем, что на протяжении сотен тысяч лет ягуар не был верховным хищником (в Южной и Центральной Америке он занимает нишу верховного хищника лишь последние 10 тыс. лет). В плейстоцене на территории Нового Света существовало, по меньшей мере, 10 видов хищников крупнее ягуара (1,12). Не будучи верховными хищниками, ягуары должны были проявлять известную гибкость, в том числе и в плане охотничьего поведения.
Предположения Хэйварда и его коллег, несмотря на всю их логическую последовательность, остаются всего лишь предположениями до тех пор, пока объективные данные не поставят точку в этом вопросе. Прежде всего, речь идет о данных изотопного анализа, данных тафономии и данных анализа механических повреждений костей потенциальных жертв.
Фабиана Мартин, изучавшая останки животных из пещер южной Патагонии, обнаружила, что останки ягуаров (Panthera onca mesembrina) очень часто находятся вместе с останками крупных травоядных животных (ламы, камелиды, милодоны, макраухении и т.д.). В данной местности наиболее вероятными конкурентами ягуара за крупную добычу представляются Smilodon populator, Arctotherium tarijense и Felis concolor (4). Плейстоценовая форма южноамериканского ягуара (Panthera onca mesembrina) отличалась очень крупными размерами, достигавшими размеров современных львиц (120-160 кг). Плейстоценовые пумы, кажется, не демонстрируют размеров больших, чем крупные современные (13). Учитывая, что и в настоящее время в местах, где ареал пумы и ягуара пересекается, ягуар чаще охотится на более крупную добычу, а пума, наоборот, представляется совершенно невероятным иное положение дел в плейстоцене, когда разница в размерах между пумой и ягуаром была еще более выразительной. Франсиско Превости и Сержио Вискаино, изучавшие размеры крупных хищников плейстоцена Аргентины, пришли к выводу, что масса Arctotherium tarijense колебалась в интервале 102-189 кг (средняя масса – 139 кг), что лишь немногим больше массы плейстоценовых аргентинских ягуаров (интервал 95-137 кг, средняя масса - 119 кг). Учитывая, что масса патагонских плейстоценовых ягуаров была большей, чем у аргентинских (4,344), разница с Arctotherium tarijense перестает выглядеть существенной. Как показывают расчеты Превости и Вискаино, средний размер типичной добычи у плейстоценовых ягуаров (135 кг) и арктотериев (100 кг) был практически одинаковым (3,tabl.2). Это и не удивительно, учитывая схожие размеры хищников. Однако Arctotherium tarijense, как и большинство медведей, был всеядным и, вероятно, охотился гораздо реже ягуара, являющегося гиперхищником. Более того, если проецировать взаимоотношения «тигр – бурый медведь» в уссурийской тайге на взаимоотношения «Panthera onca mesembrina - Arctotherium tarijense» в аргентинской пампе эпохи плейстоцена, то с учетом весового паритета южноамериканских хищников, можно предположить, что арктотерии, скорее всего, стремились избегать встреч с соразмерной им крупной кошкой. Известно также, что медведи очень редко аккумулируют кости. Следовательно, едва ли кости травоядных, найденные в патагонских пещерах, являются остатками добычи арктотериев. Наоборот, подобная аккумуляция свойственна крупным кошкам. Исследование формы и размера отметок от зубов хищников на костях животных, найденных в Lago Sofia, показало, что они были оставлены плотоядным более крупным, чем современная пума. Учитывая, что в этой пещере были найдены останки минимум трех особей Panthera onca mesembrina (включая одного детеныша) можно с очень высокой долей вероятности утверждать, что данная пещера служила логовом ягуаров, куда они и носили свою добычу. В Lago Sofia отметки от зубов крупного хищника содержат кости следующих животных: Lama sp., Mylodon darwini, Hippidion saldiasi. Сложно сказать охотились ли ягуары на взрослых Mylodon darwini, весом около 1000 кг (14). Но детеныши и юные особи были им вполне по зубам, о чем и свидетельствует найденная в Lago Sofia часть плечевой кости юной особи со следами зубов хищника (4,347).
Двухсоткилограммовые лошади кажутся вполне нормальной добычей для 150 кг кошки. В Milodon cave кости Hippidion saldiasi и милодонтин также несут отметки от зубов кошки, более крупной, чем современная пума и схожие с отметками современного ягуара (4,350). Фабиана Мартин приходит к выводу, что и в Milodon cave отметки на костях принадлежат крупной пантере (4,355). Ситуация в Chingues cave напоминает таковую в Lago Sofia. Здесь также найдены не только останки крупной особи Panthera onca mesembrina, но и нижняя челюсть юной особи, что позволяет рассматривать и эту пещеру как логово, куда ягуары приносили добычу. В этом месте были найдены и останки медведей, но размер и форма отметок на костях лошадей (Hippidion saldiasi) соответствуют зубам крупной кошки, а не медведя (4,356). Судя по морфологии, ягуарам принадлежат и отметки на костях камелид в Puma cave (4,358). Кости милодонов (как взрослых, так и юных) и лошадей со следами, оставленными крупными фелидами найдены в Fell cave (4,358-360). В Arroyos кости лошадей имеют отметки от зубов как крупных фелид, так и мелких канид.
Вопрос о том, могли ли, по крайней мере, некоторые из отметок на костях, найденных в патагонских пещерах, принадлежать Smilodon populator является дискуссионным. Традиционно считалось, что саблезубые избегали контактировать с костями, предпочитая срезать мясо с помощью высокоспециализированных хищнических зубов. Однако кости, собранные гомотериями в Friesenhan cave, демонстрируют отметки от зубов. Вместе с тем, исследователи, изучавшие отметки на костях, оставленные гомотериями, пришли к выводу, что «отметки от зубов редко проходят дальше кортикального слоя» (5,529). Кроме того, находки Smilodon populator в патагонских пещерах не столь многочисленны как Panthera onca mesembrina и у нас нет оснований рассматривать данные пещеры как логово смилодонов. Все это позволяет рассматривать именно Panthera onca mesembrina как главного ответственного за сбор костей в пещерах и позволяет сделать вывод, что плейстоценовые ягуары действительно предпочитали более крупную добычу, включающую лошадей, лам, камелид и молодых милодонтид. Возможно, что в отдельных случаях объектами охоты Panthera onca mesembrina могли становиться и взрослые Mylodon darwini, например старые или больные особи.
Как уже говорилось, главными конкурентами Panthera onca mesembrina были Smilodon populator, Arctotherium tarijense и Felis concolor. Анализ южноамериканских местонахождений плейстоценовых ягуаров показывает, что чаще всего им приходилось сосуществовать с пумой и смилодоном. Находки Arctotherium tarijense, кажется, встречаются чуть реже, но, тем не менее, достаточно часто, чтобы рассматривать этого медведя как типичного конкурента Panthera onca mesembrina. Осознавая всю отрывочность и неполноту информации по местонахождениям ягуаров в Южной Америке, все же отметим, что находки Felis concolor сопутствовали находкам ягуара в 15 случаях, находки Smilodon populator – в 14, а находки представителей рода Arctotherium – в 13.
Учитывая, что разница в размерах между пумой и ягуаром в плейстоцене была больше, чем сейчас, вполне обоснованным кажется предположение, что в плейстоцене пума старалась в максимальной степени избегать встреч с Panthera onca mesembrina. Хорошим аналогом здесь выступает пара «лев – леопард» в Африке. Одним из механизмов снижения межвидовой конкуренции является темпоральная сегрегация, когда конкурирующие виды охотятся в разное время суток. Подобная сегрегация пумы и ягуара отмечена в сухих лесах южной Боливии (33). При этом исследователи пришли к выводу, что такого рода сегрегация была вызвана не активностью жертв этих кошек, а их стремлением избежать возможных встреч. Вероятно, подобные механизмы действовали и в прошлом. Конкуренция между пумой и ягуаром, снижалась и за счет разницы в предпочитаемой добыче. Безусловно, пуме приходилось чаще довольствоваться более мелкой добычей. Уход пумы в сторону более мелкой добычи в местах сосуществования с ягуаром неоднократно фиксировался исследователями. Например, в Венесуэле пума и ягуар охотятся на ошейникового пекари (Pecari tajacu), но пума убивает преимущественно молодых особей (6,169; 11). И хотя у современной пумы и ягуара в местах их сосуществования рацион перекрывается на 70-45 %, тенденция к охоте на более мелкую, по сравнению с ягуаром, добычу у пумы выражена отчетливо. В плейстоцене зона перекрытия была, вероятно, ниже именно за счет того, что ягуары были крупнее и предпочитали более крупную добычу. При охоте на крупную добычу ягуары, благодаря своим большим размерам, имели преимущество перед пумой. Это хорошо заметно, например, в случае с Mylodon darwini. Если ягуары достаточно регулярно охотились на молодых, а порой и на взрослых животных, то пума вряд ли рассматривала милодонов в качестве объектов охоты. Будучи почти вдвое легче ягуара, обладая менее мощными клыками и челюстями, она вряд ли смогла бы с минимальными рисками справиться даже с молодым животным (речь идет не о детенышах, а о юных животных еще не достигших полной зрелости). С другой стороны, более легкая и быстрая пума должна была иметь преимущество при охоте на лам и некрупных лошадей. Гуанако (Lama guanicoe) и викунья (Vicugna vicugna) и сейчас являются главными объектами охоты пум в Патагонии (7,91). Вместе с тем, установлено, что, по крайней мере, в Патагонии ягуары регулярно потребляли Hippidion saldiasi. Т.е. конкуренция за эту добычу могла быть более острой. И вполне вероятно, что пуме нередко приходилось оставлять ягуарам свою добычу. Это кажется тем более вероятным, что у пумы нет привычки, свойственной леопардам, затаскивать свою добычу на дерево, спасая ее от надоедливых падальщиков и крупных хищников вроде львов и гиен. В Йеллоустоуне и национальном парке Бэнфф (Banff) волки посещали остатки трапезы пум в 22 и 33 % удачных охот кошки, а прямо отбирали добычу – в 6 и в 14 % соответственно. В центральном Айдахо и национальном парке Гласьер (Glacier) волки посещали остатки трапез пум в 18 и 20 % случаев успешной охоты (6,165). Медведи (Ursus americanus и Ursus arctos) посещали остатки трапез пум в национальном парке Гласьер и в Йеллоустоуне в 15 и 33 % (6,166). Эти примеры показывают, что пумы способны обеспечивать пропитание не только себе, но и отчасти своим более сильным конкурентам. Возможно, что в Южной Америке роль волков и бурых медведей играли ягуары и арктотерии. Вполне вероятно, что в местах сосуществования Panthera onca mesembrina и Felis concolor имела место и пространственная сегрегация. Тот факт, что останки обоих хищников часто находят в одних местах, не является препятствием для использования такого механизма снижения межвидовой конкуренции. Современные исследователи (6,169) отмечают, что в местах сосуществования ягуары чаще придерживаются мест около воды и лесных зарослей (более 500 м от кромки леса). Пумы же в таких случаях стараются придерживаться сухих мест (10,4). Трудно сказать придерживались ли плейстоценовые ягуары тех же экологических ниш, что и их современные родственники. Но учитывая тягу современных ягуаров к воде (едва ли это является совсем недавним приобретением) и широкий спектр возможностей для охоты около водопоев, кажется вероятным, что и в плейстоцене ягуары чаще могли придерживаться подобных мест обитания.
Относительно Arctotherium tarijense предполагается, что он предпочитал открытые пространства (8,226). Это обстоятельство могло способствовать пространственной сегрегации хищников. Несмотря на свои относительно крупные размеры (кондилобазальная длина черепа – 326 мм), арктотерий едва ли представлял большую опасность для взрослых особей Panthera onca mesembrina. Не превосходя размерами крупного гималайского медведя, он сам рисковал стать добычей крупной кошки. По крайней мере, это представляется вполне справедливым в отношении молодых особей и самок арктотериев, которые действительно могли стать жертвами ягуара, размером с уссурийскую тигрицу. Снижению межвидовой конкуренции в данном случае способствовала и всеядность арктотерия. На протяжении плейстоцена размеры арктотериев уменьшались. Если раннеплейстоценовые Arctotherium angustidens были гигантами, самыми крупными наземными представителями отряда Carnivora, то их позднеплейстоценовые потомки, в том числе и Arctotherium tarijense, существенно измельчали. Леонардо Сойбелзон и Борхиа Фигуэйридо, на основании изучения краниодентальной морфологии арктотериев, пришли к выводу, что диета Arctotherium tarijense уже включала большой процент растительной пищи (Arctotherium angustidens были гораздо более плотоядными и активными хищниками). Уклон в сторону всеядности, а у некоторых видов арктотериев и в сторону преимущественно растительной пищи Сойбелзон и Фигуэйридо объясняет широким распространением в среднем плейстоцене большого числа гиперхищников, в том числе и ягуаров (9,218). Если на территории Чили с ягуарами сосуществовали относительно крупные Arctotherium tarijense, то в Бразилии им компанию составляли Arctotherium wingei (некоторые исследователи рассматривают его как синоним Arctotherium vetustum) . Это были самые мелкие представители рода, с основной длиной черепа 203-215 мм (8,225). Учитывая, что средний вес более крупного Arctotherium bonaeriense Превости и Вискаино определяли в 110 кг, едва ли его меньший родственник весил больше центнера. Сойбелзон и Фигуэйридо полагают, что этот медведь был почти полностью травоядным (9,221). С учетом данных обстоятельств, Arctotherium wingei следует рассматривать не столько как конкурента Panthera onca mesembrina, сколько как его потенциальную добычу. Современный очковый медведь, Tremarctos ornatus, имеет почти такие же размеры как Arctotherium wingei и, будучи всеядным, питается преимущественно растительной пищей. В настоящее время ареалы Tremarctos ornatus и Panthera onca практически не пересекаются. Вероятно, именно этим объясняется тот факт, что в работах, посвященных ягуару, очковый медведь не рассматривается ни как конкурент, ни как потенциальная или реальная жертва (10).
Гораздо сложнее дать характеристику межвидовому взаимодействию в паре Panthera onca mesembrina - Smilodon populator. В настоящее время нет аналогов крупной саблезубой кошки. С одной стороны все выглядит предельно просто. Смилодон был гораздо крупнее плейстоценового ягуара. Масса Smilodon populator в отдельных случаях могла превышать 400 кг, но даже средняя масса была вдвое больше, чем у крупных особей Panthera onca mesembrina (12). Более того, применительно к североамериканскому Smilodon fatalis некоторыми исследователями высказывались весьма аргументированные предположения о возможной социальности этой саблезубой кошки. Относительно Smilodon populator сказать что-то по данному вопросу гораздо сложнее. С одной стороны у нас нет здесь тех аргументов, которые использовали сторонники социальности Smilodon fatalis в отстаивании своего предположения, а с другой стороны, учитывая близкое родство этих видов, мы не можем полностью ее исключить. Но в любом случае, ягуар должен был избегать встреч со смилодоном. Современные крупные кошки, обычно, не терпят конкурентов. Если встречи избежать не удалось, то представители более сильных видов, почти всегда, стараются убить представителей более слабых. Самый яркий пример такой межвидовой конкуренции мы находим в африканской саванне, где львы при малейшей возможности стараются убить леопардов или гиен. У нас нет никаких оснований подозревать Smilodon populator в большей толерантности к своим конкурентам. Очевидно, что Smilodon populator охотился на более крупную добычу. Превости и Вискаино полагают, что типичный размер его добычи – 764 кг, а вес максимальной добычи мог достигать почти 2 т (3,409). Учитывая размеры, и морфологию Smilodon populator, можно предположить, что он мог охотиться на крупных и не очень быстрых животных вроде токсодонов, макраухений, молодых хоботных и милодонтин. Т.е. Smilodon populator охотился на животных, которые в обычных ситуациях были недоступны для Panthera onca mesembrina. Едва ли смилодоны пренебрегали и более мелкой добычей – лошадьми, камелидами, но их массивное тело с укороченными дистальными частями конечностей явно не благоприятствовало регулярной охоте на таких быстрых животных. Наоборот, вся их посткраниальная морфология наводит на мысль о специализации на подавлении сопротивления очень крупной добычи, чья масса в 2-5 раз превышает массу самого хищника. Таким образом, обилие очень крупных травоядных при почти полном отсутствии конкурентов в данном размерном классе (после вымирания к среднему плейстоцену Arctotherium angustidens смилодон оказался единственным хищником способным регулярно охотиться на таких крупных животных) создавало Smilodon populator весьма комфортные условия и избавляло его от острой конкуренции с Panthera onca mesembrina. В новой работе (в печати), посвященной палеоэкологии Smilodon populator в аргентинской пампе, сделанные выше предположения получают подтверждение в виде результатов изотопного анализа (29). Общий вывод исследователей заключается в том, что Smilodon populator, живший 15 тыс. лет назад в Пампе, потреблял преимущественно крупную добычу, обитавшую на открытых пространствах, таких как макруахении и гигантские ленивцы. При этом его главными конкурентами оказывались крупные каниды рода Protocyon, а не ягуар, предпочитавший охотиться на добычу меньшего размера. Ягуар демонстрирует более низкие по сравнению со смилодоном значения по δ15N, указывающие, что его диета включала больше жертв с низким показателем по δ15N, возможно лошадей. Protocyon, будучи намного меньше Smilodon populator, не был падальщиком, подбиравшим объедки от трапезы саблезубой кошки. Его морфология указывает на сходство с современными гиеновыми собаками, известными как активные охотники. Следовательно, в аргентинской пампе главными конкурентами смилодона был не ягуар, а крупные каниды. Изотопный анализ останков животных в патагонских пещерах также подтверждает обоснованность ранее сделанных выводов. В частности, он показывает, что Panthera onca mesembrina употреблял в пищу преимущественно Hippidion saldiasi и Lama gracilis (15). Пума, вероятно, охотилась на камелид и нанду. Smilodon populator, не имея четко выраженной предпочитаемой добычи, по-видимому охотился на всех крупных животных, которых мог добыть. Арктотерии (видовая принадлежность не указана) были всеядными, и, судя по результатам изотопного анализа, довольно часто употребляли падаль (15). Подобной всеядной диеты придерживался и Dusicyon avus.
Палеоэкология североамериканского плейстоценового подвида Panthera onca augusta кажется куда менее изученной. Нам не известны работы, рассматривавшие особенности питания данного хищника с позиции результатов изотопного анализа или анализа механических повреждений костей его потенциальных жертв. Здесь мы скорее можем делать выводы по фаунистическим спискам и картам распространения тех или иных видов.
Вместе с тем, у нас есть все основания предполагать, что и Panthera onca augusta предпочитал большую добычу. Куртен и Андерсон обратили внимание на череп Glyptotherium texanum (F:AMNH 95737), в котором имеются два четко выраженных отверстия, «вероятно, сделанных клыками атаковавшей кошки» (16,134). Исследователи не утверждали, что глиптотерия убил именно ягуар, но такой вывод напрашивается после получения дополнительной информации. Glyptotherium texanum – самый ранний глиптодонт в Северной Америке, известный с позднего Бланко по находкам в Техасе и Аризоне. Это был самый мелкий североамериканский глиптодонт, с общей длиной около 220 см и весом в 450-550 кг. От Glyptotherium texanum произойдет более крупный Glyptotherium arizonae, широко распространившийся по всему югу США в Ирвингтонском веке. Следовательно, Glyptotherium texanum не мог быть жертвой Panthera atrox, поскольку «ужасные львы» в Америке появятся гораздо позже. Не мог он быть убит и саблезубыми кошками (гомотерий, смилодон), которые едва ли решились бы использовать свое страшное, но относительно хрупкое оружие, таким «варварским» способом. Кроме того, и сами отверстия в этом случае должны были бы иметь совсем другую форму и размер. Из крупных кошек верхнего плейстоцена остается только Panthera onca augusta. Кроме того, именно такой способ убийства (укус в сам череп) является «визитной карточкой» современного ягуара. Современные ягуары очень часто прибегают к такому способу умерщвления своих жертв, тогда как другие пантерины предпочитают завершать дело укусом в горло или в шею. Кажется, что современные ягуары, известные не только своим укусом в череп, но и прокалыванием панцирей черепах, не очень боятся сломать себе клыки. Возможно, это объясняется тем, что относительно своих размеров ягуары имеют очень мощные клыки и обладают очень большой силой укуса. В настоящее время, исследователи рассматривают усиление защиты глиптодонтов на протяжении плейстоцена как реакцию на активность крупных хищников, с которыми они ранее не сталкивались (17,8-9). Вместе с тем, у нас нет оснований утверждать, что глиптодонты были обычной добычей плейстоценового ягуара. Во-первых, ареал обитания Panthera onca augusta был шире, чем Glyptotherium, и во многих местах обитания ягуаров глиптотериев просто не было. Глиптодонты предпочитали теплую и влажную среду обитания. Во-вторых, сами глиптодонты, как уже отмечалось, довольно быстро эволюционировали в плане усиления своей защиты (17) и увеличения размеров. В частности, пришедший на смену Glyptotherium texanum аризонский глиптотерий (Glyptotherium arizonae) был уже более крупным животным, длиной 3 м и весом около тонны (16,134). Ягуары в Северной Америке на протяжении среднего – позднего плейстоцена, наоборот, уменьшались в размерах (18,14). К сожалению, мы не имеем подробного списка фауны всех местонахождений североамериканского ягуара. Но если опираться на мексиканские местонахождения, к которым есть списки, то мы не найдем указаний на их сосуществование ни в Colorado River Delta, ни в El Golfo, ни в Aguascalientes.
Но если глиптодонты были скорее случайными жертвами, то кто же составлял список основных жертв Panthera onca augusta? В работе Мартина и Шульцов есть очень интересная карта, показывающая места находок тапира, длинноносого пекари и ягуара (19,fig.3).

На карте мы видим, что во многих случаях места находок ягуара и его потенциальных жертв перекрываются. Добавим, что и на территории Мексики известны местонахождения, доказывающие сосуществование ягуаров и тапиров на одной территории (El Golfo, Colorado River Delta). Вполне вероятно, что более крупные, чем современные, ягуары плейстоцена гораздо чаще охотились на тех же тапиров и пекари. Размер тапиров и длинноносых пекари кажется вполне оптимальным для типичной добычи 120 кг кошки. Если говорить о пекари, то, разумеется, нельзя ограничиваться только длинноносым пекари, Mylohyus nasutus. Тут скорее уместно вести речь о представителях рода Mylohyus в целом. Доказательством этому служат находки ягуара и других видов пекари из этого рода в Cumberland Cave, и в Port Kennedy Cave. Очень часто останки Panthera onca augusta находят вместе с останками представителей семейства пекариевых из рода Platygonus. Такого рода местонахождения есть в Аризоне, Пенсильвании, Мэриленде, Орегоне, Техасе, Индиане. Находки позднеплейстоценовых Platygonus compressus известны даже с Юкона (22). Пекари из рода Platygonus, как и их родственники из рода Mylohyus, не были особо крупными животными. В частности вес Platygonus compressus (останки этого вида пекари были найдены вместе с останками ягуара в Laubach Cave, Техас, США) был оценен в 75 кг (20,163). Вполне вероятно, что во многих популяциях Panthera onca augusta именно пекари составляли основу его рациона. Данный вывод заставляет нас усомниться в высказываемом некоторыми учеными предположении, что современный ягуар избегает охоты на пекари, боясь получить раны. Плейстоценовые ягуары охотились на них, даже при куда большем выборе потенциальной крупной добычи. Возможное возражение, что Panthera onca augusta был крупнее современного ягуара, парируется тем, что масса большинства видов современных пекари не достигает и 50 кг (21). Широкий ареал обитания, высокая плодовитость и умеренно крупные размеры – все это позволяет рассматривать пекари как одну из типичных жертв Panthera onca augusta.
Часто (от Орегона до Флориды и от Калифорнии до Мэриленда) в находки североамериканских ягуаров сопровождаются находками эквид. Учитывая, что эквиды составляли значительную часть рациона Panthera onca mesembrina, кажется уместным предположение, что и североамериканские плейстоценовые ягуары могли активно охотиться на лошадей. Чуть реже, но тоже довольно часто, в местонахождениях ягуаров находят останки цервид из рода Odocoileus. Виргинский олень (Odocoileus virginianus), находки которого сопровождают находки ягуара в целом ряде американских штатов (Пенсильвания, Техас, Арканзас), пережил вымирание позднего плейстоцена. Это средних размеров олень, массой до 150 кг. Учитывая огромный ареал обитания этого оленя, как в прошлом, так и в настоящем, и тот факт, что места его находок часто совпадают с местами находок ягуара, данный вид оленя, равно как и оленей из рода Odocoileus, также следует отнести к типичной добыче Panthera onca augusta. Камелиды, и в частности представители рода Hemiauchenia, также довольно часто являются компонентом сопутствующей фауны и могут рассматриваться, как вероятная добыча ягуаров. Hemiauchenia macrocephala (чаще всего, когда видовая принадлежность определена и указана, речь именно о ней) была крупнее современных лам, имела более длинные конечности и удлиненные метаподии, что указывает на курсориальность и адаптацию к открытым ландшафтам (26,43). Вероятно ирвингтонским ягуарам, имевшим по сравнению со своими ранчолабрейскими потомками более длинные конечности и пропорционально более длинные дистальные части конечностей (18,14), было легче охотиться на этих животных.
Охота одиночных Panthera onca augusta на молодых хоботных представляется маловероятной. В современной Африке максимальному риску нападения львов подвергаются не маленькие детеныши, которые держатся около взрослых самок, а уже подросшие, но не достигшие полной зрелости, особи, которые позволяют себе отделяться от стада. Но даже в этом случае, в убийстве такого слона-подростка, как правило, принимает участие несколько членов львиного прайда. Североамериканские плейстоценовые ягуары, размеры которых не превышали размеры современных львиц, вряд ли были способны в одиночку справиться с такой задачей. Поэтому, юные особи хоботных мог ли быть лишь редкими, случайными жертвами Panthera onca augusta.
Крупные плейстоценовые бизоны (Bison latifrons и Bison antiques), останки которых порой сопровождают останки Panthera onca augusta, едва ли были их добычей. Бизоны появились в Северной Америке довольно поздно, и рассматриваются как типичные представители ранчолабрейской фауны. Плейстоценовые бизоны, высотой более 2 м, были огромными животными, заметно превосходившими своих современных потомков размерами и силой. Справиться в одиночку с бизонами весом 800-1500 кг Panthera onca augusta было практически не реально. Отбить от стада детеныша в одиночку - не менее сложная задача. Кроме того, вслед за бизонами в Новый Свет пришли гигантские львы, Panthera atrox, весившие в два с половиной раза больше плейстоценовых ягуаров. Конкурировать с крупными, социальными и курсориальными львами на равнинах прерий ягуары не могли. Кроме того, как показывают данные изотопного анализа, бизоны были излюбленной добычей крупных саблезубых кошек Smilodon fatalis (23,211). Как и «ужасные львы» смилодоны могли быть социальными животными. Во всяком случае, аргументы в пользу этой гипотезы имеются, и хотя она принимается далеко не всеми исследователями, мы не можем исключать социальность Smilodon fatalis. Но даже одиночный смилодон, весивший 250-300 кг, мог представлять серьезную угрозу для 120 кг ягуара. Таких встреч ему желательно было избегать.
Но на бизонов, как показывают данные того же изотопного анализа, весьма активно охотились не только львы и саблезубые кошки, но и «ужасные волки», Canis dirus. Если относительно размеров прайдов Panthera atrox исследователи осторожно высказываются в пользу небольшого числа особей (т.е. социальность львов практически никем не отрицается, но допускается меньший размер прайда, чем у современных львов), а относительно социальности Smilodon fatalis нет единого мнения, то социальность Canis dirus сомнений не вызывает. Это были крупные волки, весившие 60-68 кг (24,211), обладавшие более крупными, по сравнению с Canis lupus, зубами и более мощными челюстями. Стаи таких волков на протяжении тысяч лет успешно конкурировали даже со смилодонами и «ужасными львами». При встрече с ними у ягуара, скорей всего, было мало шансов отстоять свою добычу. На первый взгляд, в случае с парой «Panthera onca augusta - Canis dirus» (останки обоих хищников находили в одних и тех же местонахождениях Пенсильвании, Калифорнии, Орегона, Южной Каролины, Техаса, Индианы, Нью-Мексико) напрашивается аналогия «Panthera tigris altaica - Canis lupus chanco». Аналогия отчасти обоснована тем, что и в том и в другом случае речь идет о межвидовой конкуренции крупной одиночной пантеры с волками. Размеры Panthera onca augusta вполне укладываются в размеры самок уссурийских тигров. Однако волки, обитающие в Уссурийской тайге, относятся к мелкому подвиду. Масса самцов Canis lupus chanco не превышает 37 кг (25,31). Кроме того, в уссурийской тайге волкам приходится иметь дело не только с тигрицами, но и с тиграми, которые уже заметно крупнее плейстоценовых ягуаров. «Ужасные волки» были вдвое крупнее Canis lupus chanco. И если в Уссурийской тайге волки всегда отступают перед тиграми, не пытаясь оспаривать право на добычу даже взрослых тигриц, то в плейстоцене Северной Америки ситуация могла быть совсем иной. Стая Canis dirus не только могла отстоять свою добычу от ягуара, но и сама могла лишить его добычи, окружая и нападая с разных сторон. В такой ситуации для любой одиночной кошки правильной стратегией поведения является отступление, а не борьба, чреватая серьезными травмами, а то и смертельным исходом. Таким образом, конкуренция за бизонов между Panthera atrox, Smilodon fatalis и Canis dirus была очень острой. К этой троице необходимо добавить и гигантского короткомордого медведя, который, предпочитая открытые пространства, всегда был готов отнять добычу у более слабых конкурентов. В данной ситуации Panthera onca augusta было гораздо проще переключиться на другую доступную добычу, чем пытаться конкурировать с заведомо более сильными соперниками за бизонов, взрослые особи которых были ему просто не по зубам.
Как и в Южной Америке, молодые милодонтины вполне могли быть объектом охоты ягуаров. Но едва ли они по популярности могли конкурировать с пекари, эквидами, камелидами и оленями. Хотя бы потому, что в случае с большинством видов милодонов объектами охоты могли стать только юные особи, не достигшие полной зрелости. Paramylodon harlani (=Mylodon harlani= Glossotherium harlani), останки которого были найдены вместе с останками ягуара в целом ряде мест на территории США (Port Kennedy Cave (Пенсильвания), Rancho La Brea (Калифорния), Rome Beds (Орегон), Fossil Lake (Орегон)), был крупным животным, высотой 130 см (в спине на четырех конечностях) и весом в 1,5 т. (26,52; 27,39). Он имел мощные передние конечности, вооруженные большими, сильными когтями. Дермальные костные бляшки в толще шкуры обеспечивали дополнительную защиту, особенно в области шеи и плечевого пояса (26,51). Энергетические затраты, и, что более существенно, риски получить смертельную травму должны были удерживать Panthera onca augusta от охоты на такого мощного, хорошо защищенного зверя.
Некоторые нотротериопсы, вроде Nothrotheriops shastensis, вполне могли быть добычей ягуаров. Останки этих нотротериопсов и Panthera onca augusta найдены в Rancho La Brea (Калифорния) и в Dry Cave (Нью-Мексико). Учитывая их размеры (вес 135-180 кг), они выглядят вполне подходящей добычей для этой кошки.
На протяжении плейстоцена ситуация с межвидовой конкуренцией для Panthera onca augusta только ухудшалась. В раннем плейстоцене главными конкурентами были Miracinonyx inexpectatus, Smilodon gracilis, Homotherium serum, Felis concolor, Arctodus pristimus. В среднем плейстоцене ситуация начинает заметно ухудшаться. На смену 100 кг Smilodon gracilis приходит 280 кг Smilodon fatalis, на смену Arctodus pristimus – гигантский Arctodus simus, весом более 800 кг (28). В позднем плейстоцене (ранчолабрейский век) конкуренция обостряется еще сильнее. Вслед за бизонами в Америку прибыли «ужасные львы», Panthera atrox. Место Canis armbrusteri (размером с Canis lupus) окончательно занимает более крупный Canis dirus (36). Конкуренция на открытых пространствах ужесточается до предела. И в этой борьбе далеко не самая крупная и не самая быстрая одиночная кошка оказывается «слабым звеном».
Рассмотрим ситуацию с межвидовой конкуренцией в Северной Америке в плейстоцене чуть более подробно. Как уже говорилось, в раннем плейстоцене основными конкурентами Panthera onca augusta были Miracinonyx inexpectatus, Smilodon gracilis, Homotherium serum, Felis concolor, Arctodus pristimus. Мирациноникс был крупной кошкой, представляющей пумо-гепардовскую линию фелид. Пропорции позвоночника, морфологические особенности и пропорции конечностей, рассматриваемые сквозь призму адаптаций к быстрому бегу, указывают на то, что Miracinonyx inexpectatus занимал в этом плане промежуточное положение между пумой и гепардом (37). Miracinonyx inexpectatus, был крупнее современного гепарда и весил примерно 70 кг (37,452). С учетом того, что кости конечностей Miracinonyx inexpectatus были заметно крупнее, чем у современной пумы, возможно, эти кошки весили даже больше. Останки Miracinonyx inexpectatus и Panthera onca augusta, датированные ранним – средним плейстоценом, часто находят вместе (Hamilton Cave, Port Kennedy Cave, и Conard Fissure). В этот период, по-видимому, острой конкуренции этим двум видам удавалось избегать за счет пространственной сегрегации. Мирацинониксы, были явно адаптированы к скоростному преследованию на открытых пространствах, хотя еще и не достигли в этом плане специализации гепарда. Как отмечали Антон и Тёрнер, эта кошка была «более быстрой, чем пума, но более мощной и лучше экипированной для лазанья по деревьям, чем гепард» (38,90). Ирвингтонские Panthera onca augusta, хотя и имели удлиненные дистальные части конечностей (адаптация к скоростному бегу) в скорости явно проигрывали мирацинониксам. Вероятно, Panthera onca augusta предпочитал держаться более закрытых ландшафтов, где недостаток скорости можно было компенсировать подкрадыванием на более короткую дистанцию. С учетом того, что ягуары были все же крупнее и заметно мощнее, логично предположить, что Miracinonyx inexpectatus в большей степени был заинтересован в избегании встреч со своим конкурентом. Возможно, в чем-то отношения этих двух кошек напоминали отношения пары «гепард – леопард» в современной Африке. Гепарды, по массе не сильно уступающие леопардам, тем не менее, почти всегда вынуждены уступать им свою добычу. Ведь в случае прямого столкновения, у этой кошки, пожертвовавшей ради скорости силой и мощью, будет мало шансов устоять против леопарда, заметно превосходящего гепарда не только мощью мускулатуры, но и длиной клыков и силой укуса. Если это так, то вполне вероятна и темпоральная сегрегация между этими видами. Известно, что современные гепарды охотятся преимущественно днем, в то время, когда полуденная жара заставляет львов и гиен искать убежища в тени. В современной Африке леопарды сами стараются нечасто выходить на открытые пространства, предпочитая держаться кромки леса, где ближайшее дерево уже может служить спасением от львов. У Panthera onca augusta в раннем и среднем плейстоцене не было необходимости вести себя столь осторожно. Поэтому вероятность появления ягуара на открытых пространствах была выше, что в свою очередь увеличивает вероятность наличия темпоральной сегрегации между этими двумя хищниками. Снижение межвидовой конкуренции возможно и за счет того, что хищники охотились на разную добычу. Если ягуары охотились на пекари (представители рода Mylohyus были найдены и в Port Kennedy Cave и в Conard Fissure), то мирацинониксы могли предпочитать эквид или оленей.
Homotherium serum представлял собой крупную саблезубую кошку размером с современного льва. Гомотерий, как плейстоценовый ягуар, был очень широко распространен на всем североамериканском континенте: от Аляски до Техаса и от Калифорнии до Флориды. Находки ягуара и гомотерия зафиксированы в Калифорния (Rancho La Brea и Irvington), Флориде (Reddick I) и, возможно в Техасе (Friesenhahn Cave). Homotherium serum традиционно рассматривался как охотник на очень крупную добычу, в частности на молодых хоботных. В этом исследователей убеждали найденные в пещере Фризенхахн, служившей логовом гомотерию, останки по меньшей мере 34 колумбийских мамонтов (5,fig.1). Кроме колумбийских мамонтов в этой же пещере были найдены останки американского мастодонта. Однако последние исследования показывают, что гомотерий не был узкоспециализированным хищником. Данные изотопного анализа показывают, что позднеплейстоценовые гомотерии Берингии потребляли преимущественно яков (Bos mutus), дополняя свой рацион северными оленями, бизонами и овцебыками (41,60). Следовательно, было бы ошибочно исключать его из числа конкурентов ягуара. Вместе с тем, морфология гомотерия позволяет рассматривать его как курсориального хищника открытых пространств. Следовательно, конкуренция между этими двумя видами снижалась за счет пространственной сегрегации. Другим механизмом снижения была охота на разную добычу. Хотя, как показывают последние исследования, гомотерий успешно охотился и на животных, которых нельзя отнести к особо крупным (северный олень), основу его рациона все же составляли животные, на которых вряд ли охотился ягуар (хоботные в Техасе, яки и бизоны в Берингии) в силу своих меньших размеров. К моменту прибытия ягуара в Новый Свет гомотерии были самыми крупными кошками североамериканского континента. Позже, они уступят это первенство смилодонам и «ужасным львам». Можно предположить, что конкуренция между Panthera onca augusta и Homotherium serum была все же не очень острой, и гомотерий не был для ягуаров смертельно опасным конкурентом, какими стали американские львы. Если Panthera atrox по массе превосходил ягуара в два с половиной раза, то гомотерий был тяжелее его всего на 50 %. Если проводить аналогии с современными хищниками, паре «Panthera onca augusta и Homotherium serum» гораздо больше соответствует пара «пума и ягуар», чем пара «леопард – лев». Это означает, что в случае встречи, например, самки гомотерия с самцом ягуара, последнему, скорее всего, удалось бы отстоять свою добычу и вообще, вряд ли пришлось спасаться бегством. Даже в случае встречи с самцом гомотерия, ягуар, скорее всего, мог не опасаться за свою жизнь. Другое дело, что он мог лишиться добычи или вынужден был бы уйти с данной территории. Скорее всего, гомотерий довольствовался бы этим. При всей нетерпимости к конкурентам вступать в смертельный бой с соперником, который не так уж сильно уступает тебе размером и силой довольно рискованное занятие. Таким образом, степень опасности для ягуара со стороны гомотерия была существенно ниже, чем со стороны Panthera atrox.
Другой конкурент, Smilodon gracilis,был почти соразмерной Panthera onca augusta саблезубой кошкой, весом 70-120 кг. Даже если в среднем Smilodon gracilis и был немного меньше ирвингтонского ягуара, едва ли это имело решающее значение. В отличие от грацильного мирациноникса, Smilodon gracilis был очень мощной кошкой, крепкого телосложения. Обладая, как и все смилодонтины, очень сильными передними конечностями, вооруженными длинными когтями и длинными клыками Smilodon gracilis представлял собой весьма серьезного соперника. О доминировании со стороны ягуара здесь говорить едва ли уместно. Скорее, это были равные по силе соперники. Межвидовая конкуренция в этом случае вероятнее всего снижалась за счет пространственной сегрегации. По-видимому, Smilodon gracilis предпочитал закрытые ландшафты в виде густых зарослей, позволявших подкрасться к добыче на минимальное расстояние. Пропорции конечностей смилодонов не позволяют видеть в них хороших бегунов. И хотя Smilodon gracilis по части укорачивания дистальных отделов конечностей было еще далеко до Smilodon populator, тенденция к примату мощи над скоростью была выражена уже весьма отчетливо. Кроме того, особенности строения отдельных костей позволили А.Берте предположить, что Smilodon gracilis был адаптирован для жизни в лесу (39,41). Вместе с тем, было бы ошибочно полагать, что Smilodon gracilis не покидали лесных чащоб. Один из самых известных экземпляров этой саблезубой кошки из Inglis IA (Флорида) как раз и доказывает возможность существования данного вида на открытых пространствах. Вполне вероятно, что именно соседство с крупными ягуарами вынудило смилодонов взять курс на увеличение размеров, что позволило перейти в другой размерный класс и начать охотиться на уже более крупную добычу. По-видимому, этот процесс завершается уже в среднем плейстоцене, с появлением крупной формы Smilodon fatalis.
В случае конкуренции со Smilodon fatalis с одной стороны есть предельная ясность, а с другой стороны – значительная неопределенность. Понятно, что Panthera onca augusta не могла тягаться с 250-300 кг саблезубой кошкой. Даже с одной. Вместе с тем, эти смилодоны, по-видимому, предпочитали те же самые закрытые и полузакрытые ландшафты, что и ягуары. Во всяком случае, именно к такому выводу подталкивает многолетние изучение самого известного местонахождения саблезубых кошек Ранчо Ла Бреа. Возможный вариант снижения внутривидовой конкуренции в данном случае – охота на разную добычу. По данным изотопного анализа излюбленной добычей Smilodon fatalis в Ранчо Ла Бреа были Bison antiques Camelops hesternus (23,211). К сожалению, в данном случае мы не можем сказать что-либо конкретное о питании ягуара. Panthera onca augusta, хотя и отмечена в Ранчо Ла Бреа, была уже довольно редким хищником в этих краях в Ранчолабрейском веке. Но, очевидно, что ягуару приходилось охотиться на менее крупную добычу, чем смилодону.
Появление в среднем и позднем плейстоцене таких крупных и, вероятно, социальных хищников как Panthera atrox и Smilodon fatalis не могло не отразиться на дальнейшей судьбе Panthera onca augusta. Ягуары начинают уменьшаться в размерах. Ранчолабрейские ягуары по массе примерно на 10 % меньше Ирвингтонских (18,14; 40,357). Дистальные части конечностей ранчолабрейских ягуаров укорачиваются. В частности, если длинные кости конечностей сокращаются на 5,8 %, то кости стопы – на 9,5 % (18,14). Такое укорачивание, по мнению Б. Куртена, является характеристикой современных ягуаров. Вместе с тем, это может означать, что ягуары окончательно вытесняются в закрытые ландшафты. Очевидно, что такое вытеснение в сочетании с уменьшением размеров могло вести и к изменению предпочитаемой добычи, когда место крупных животных все чаще стали занимать животные среднего и мелкого размера. Вполне вероятно, что это был достаточно длительный процесс, продолжавшийся на протяжении всего Ранчолабрейского века. В таком случае, проигрыш ягуаров в конкурентной борьбе смилодонам и «ужасным львам», в действительности обернулся их победой в борьбе за выживание с началом массового вымирания мегафауны. У ягуаров было достаточно времени на адаптацию, чего не было у тех же американских львов и саблезубых кошек.
1. Hayward M.W. et al. (2016) Prey Preferences of the Jaguar Panthera onca Reflect the Post-Pleistocene Demise of Large Prey // Frontiers in Ecology and Evolution. Vol.3, Article 148. pp. 1-19.
2. Из огромного массива литературы по данному вопросу отметим лишь следующие. Azevedo F.C.C., Verdade L.M. (2012) Predator-prey interactions: jaguar predation on caiman in a floodplain forest. // Journal of Zoology 286,200–207; Cavalcanti S.M.C., Gese E.M. (2010) Kill rates and predation patterns of jaguars (Panthera onca) in the southern Pantanal, Brazil // Journal of Mammalogy. 91, 722–736; Silveira R. et al. (2010) Depredation by Jaguars on Caimans and Importance of Reptiles in the Diet of Jaguar // Journal of Herpetology, 44(3):418-424; Novack A.J. et al. (2005) Foraging ecology of jaguar (Panthera onca) and puma (Puma concolor) in hunted and non-hunted sites within the Maya Biosphere Reserve, Guatemala // Journal of Zoology. 267, 167–178; Núñez R., et al. (2000) Food habits of jaguars and pumas in Jalisco, Mexico // Journal of Zoology. 252,373–379; Azevedo F.C.C. (2008) Food Habits and Livestock Depredation of Sympatric Jaguars and Pumas in the Iguac¸u National Park Area, South Brazil // BIOTROPICA 40(4): 494–500; Astete S. et al. Comparative Ecology of Jaguars in Brazil // CAT News Special Issue 4 - The Jaguar in Brazil; López González C. A., Miller B. J. 2002. Do jaguars (Panthera onca) depend on large prey? // Western North American Naturalist 62, 218-222.
3. Prevosti F.J., Vizcaino S.F. (2006) Paleoecology of the large carnivore guild from the late Pleistocene of Argentina // ACTA PALAEONTOLOGICA POLONICA 51 (3), pp. 407-422.
4. Martin F.M. (2008) Bone Crunching felids at the End of the Pleistocene in Fuego-Patagonia, Chile. // Journal of Taphonomy 6 (3-4): 337-372.
5. Marean C.W., Ehrhardt C.L. (1995) Paleoanthropological and paleoecological implications of the taphonomy of a sabertooth den. // Journal of Human Evolution, 29: 515–547.
6. Ruth T.K., Murphy K. Competition with Other Carnivores for Prey/In.: Cougar Ecology and Conservation. /Edit. by Maurice Hornocker and Sharon Negri.
7. Walker S., Novaro A. The World’s Southernmost Pumas in Patagonia and the Southern Andes/In.: Cougar Ecology and Conservation. /Edit. by Maurice Hornocker and Sharon Negri.
8. Барышников Г.Ф. Медвежьи (Carnivora, Ursidae). – СПб.: Наука, 2007.
9. Figueirido B., Soibelzon L.H. (2010) Inferring palaeoecology in extinct tremarctine bears (Carnivora, Ursidae) using geometric morphometrics // LETHAIA 43, pp. 209-222.
10. Seymour K. (1989) Panthera onca // Mammalian Species. No.340. pp. 1-9.
11. Azevedo F.C.C. (2008) Food Habits and Livestock Depredation of Sympatric Jaguars and Pumas in the Iguac¸u National Park Area, South Brazil // BIOTROPICA 40(4): 494–500.
12. Christiansen P., Harris J.M. (2005) Body Size of Smilodon (Mammalia: Felidae) // JOURNAL OF MORPHOLOGY 266:369–384.
13. В работе Франсиско Превости и Фабианы Мартин вес патагонской плейстоценовой пумы указан в 145 кг. См. - Prevosti F.J., Martin F.M. (2013) Paleoecology of the mammalian predator guild of Southern Patagonia during the latest Pleistocene: Ecomorphology, stable isotopes, and taphonomy // Quaternary International Volume 305, pp. 74–84. Но эта оценка, сделанная на основе площади нижнего хищнического зуба, кажется завышенной. Равно это касается и 190 и 243 кг Panthera onca mesembrina. Еще менее реалистичными выглядят оценки массы, сделанные на основе переднезаднего диаметра диафиза плечевой кости. Для пумы по этому параметру авторы в приложениях указывают 145 и 164 кг, а для ягуара – 269, 199 и 251 кг. Данные по медиолатеральной ширине диафиза большой берцовой кости дают более адекватные значения – 121 кг для пумы и 131-151 кг для ягуара. Внимательно просмотрев у К. Сеймура все метрические данные по посткраниальным останкам Panthera onca mesembrina, позволяющие дать более точную, по сравнению с зубами, оценку массы, мы не обнаружили костей, по размеру сопоставимых с костями крупного льва. Надо сказать, что Превости и Мартин, сами прекрасно понимая сомнительность некоторых своих оценок, в тексте предлагают для пумы и ягуара вес 121 и 140 кг соответственно. Но и эта более осторожная оценка может быть завышенной. Размеры верхних хищнических зубов плейстоценовых пум из Аргентины ненамного превышают максимальные значения современных пум, но при этом они заметно меньше аналогичных зубов современного ягуара – См. Seymour K.L. (1983). The Felinae (Mammalia: Felidae) from the late Pleistocene tar seeps at Talara, Peru, with a critical examination of the fossil and Recent felines of North and South America. M.Sc. thesis, University of Toronto, Toronto. Tabl.6. Размеры нижних хищнических зубов плейстоценовых пум Южной Америки не выходили за верхнюю границу значений современных пум, и находятся на уровне средних значений самцов современных ягуаров - Seymour K.L. Op. Cit. Tabl. 9 и 2. Лучевые и бедренные кости пум из Талары находятся в рамках средних значений современных пум (Tabl. 10 и 15). Локтевые и большеберцовые кости из Талары демонстрируют размеры чуть выше средних значений современных пум, не дотягивая до их максимальных показателей (Tabl. 14 и 16). Средних размеров оказываются и найденные в Таларе метакарпальные и метатарсальные кости пум (Tabl.20 и 21). Вывод Сеймура однозначен: «Все материалы, по сути, размером и морфологией идентичны исследованным экземпляром современной пумы (Tabl. 11-24)» - Seymour K.L. Op. Cit. p. 103. В Аргентине, по расчетам Превости и Вискаино, средняя масса плейстоценовой пумы была всего лишь 50 кг (3,tabl.1). Во Флориде позднеплейстоценовые пумы по размерам не отличались от современных пум Флориды, относящихся к одному из самых мелких подвидов – см. Morgan G.S., Seymour K.L. (1997) FOSSIL IIISTORY OF THE PANTHER (PUMA CONCOLOR) AND THE CHEETAH-LIKE CAT (M1RACINONYX lNEXPECTA TUS) IN FLORIDA // BULLETIN OF THE FLORIDA MUSEUM Of NATURAL HISTORY. Vol. 40 No.2, pp. 177-219. Возможно, патагонские плейстоценовые пумы действительно демонстрировали несколько большие размеры. В этом нет ничего удивительного, т.к. и ягуары из этих мест были чуть больше плейстоценовых ягуаров из аргентинской пампы. Но и в этом случае, едва ли их вес превышал 100 кг.
14. Вес в 1000 кг указан у Ф. Мартин. Другие исследователи указывают гораздо больший вес – 1600 кг (Vizcaino S.F. et al. (2006) Dental occlusal surface area in relation to body mass, food habits and other biological features in fossil xenarthrans // AMEGHINIANA (Rev. Asoc. Paleontol. Argent.) - 43 (1): 11-26.).
15. Prevosti F.J., Martin F.M. (2013) Paleoecology of the mammalian predator guild of Southern Patagonia during the latest Pleistocene: Ecomorphology, stable isotopes, and taphonomy // Quaternary International Volume 305, pp. 74–84.
16. Kurten B., Anderson E. (1980) Pleistocene Mammals of North America.
17. Zurita A.E. et al. (2010) Accessory protection structures in Glyptodon Owen (Xenarthra, Cingulata, Glyptodontidae) Structures accessoires de protection en Glyptodon Owen (Xenarthra, Cingulata, Glyptodontidae) // Annales de Paléontologie 96, pp. 1–11.
18. Kurtén B. 1973. Pleistocene jaguars in North America. Commentationes Biologicae, Societas Scientiarum Fennica 62:1-23.
19. Schultz C.B, Martin L.D., Schultz M.R. (1985). A Pleistocene jaguar from north-central Nebraska. // Transactions of the Nebraska Academy of Sciences and Affiliated Societies, 8:93-98.
20. Lambert W.D., Holling C.S. Causes of ecosystem transformation at the end of the Pleistocene: evidence from mammal body-mass distributions // Ecosystems (1998) 1: 157–175.
21. Mayer J.J., Wetzel R.M. (1986) Catagonus wagneri // Mammalian Species. No.259. pp. 1-5.
22. Beebe B.F. Pleistocene peccary, Platygonus compressus Le Conte, from Yukon Territory, Canada // Canadian Journal of Earth Sciences. Vol. 17, 1204-1209.
23. Coltrain J.B. et al. (2004) Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal southern California // Palaeogeography, Palaeoclimatology, Palaeoecology 205, pp. 199– 219.
24. Anyonge W., Roman C. (2006) New Body Mass Estimates for Canis Dirus, the Extinct Pleistocene Dire Wolf // Journal of Vertebrate Paleontology, Vol. 26, No. 1, pp. 209-212.
25. Бибиков Д.И. Волк. Происхождение, систематика, морфология, экология. М.: «Наука», 1985.
26. Stock Chester Rancho la Brea. A record of Pleistocene life in California // Natural History Museum of Los Angeles County № 37.
27. Baskin J.A., Thomas R.G. (2007) South Texas and the Great American Interchange: Gulf Coast Association of Geological Societies Transactions, v. 57, p. 37-45.
28. Christiansen P. (1999) What size were Arctodus simus and Ursus spelaeus (Carnivora: Ursidae)? // Ann. Zool .Fenn. 36:93–102.
29. Bocherens H. et al. (2016) Paleobiology of sabretooth cat Smilodon populator in the Pampean Region (Buenos Aires Province, Argentina) around the Last Glacial Maximum: Insights from carbon and nitrogen stable isotopes in bone collagen / In Press
30. Garla R. et al. (2001) Jaguar (Panthera onca) Food Habits in Atlantic Rain Forest of Southeastern Brazil // BlOTROPlCA 33(4): 691-696.
31. Daggett P.M., Henning D.R. (1974) The Jaguar in North America // American Antiquity, Vol. 39, No. 3, pp. 465-469.
32. Rabinowitz A.R. (1999) The Present Status of Jaguars (Panthera Onca) in the Southwestern United States // The Southwestern Naturalist, Vol. 44, No. 1, pp. 96-100.
33. Romero-Muñoz A. et al. (2010) Temporal separation between jaguar and puma in the dry forests of southern Bolivia // Journal of Tropical Ecology 26:303–311.
34. Diaz N.I. (2010) NEW HISTORICAL RECORDS OF THE JAGUAR (Panthera onca) IN PATAGONIA // Revista Mexicana de Mastozoología 14:23-45.
35. RODRIGUES S. et al. (2014) Late Pleistocene carnivores (Carnivora: Mammalia) from a cave sedimentary deposit in northern Brazil // Anais da Academia Brasileira de Ciências
36. Самая ранняя находка Canis dirus в Salamander Cave (Южная Дакота) датируется возрастом 252 тыс. лет - Dundas R.G. (1999) Quaternary records of the dire wolf, Canis dirus, in North and South America. // Boreas, Vol. 28, n.3, p.376.
37. Valkenburgh Blaire Van et al. (1990) The Plio-Pleistocene Cheetah-Like Cat Miracinonyx inexpectatus of North America // Journal of Vertebrate Paleontology, Vol. 10, No. 4, pp. 434-454.
38. Anton M., Turner A. The Big Cats and their fossil relatives. Columbia University Press, New York, 1997.
39. Berta, A., 1987, The sabercat Smilodon gracilis from Florida and a discussion of its relationships (Mammalia, Felidae, Smilodontini): Bulletin of the Florida State Museum, Biological Sciences, Vol. 31, No.1, p. 1-63.
40. Seymour K., 1993. Size changes in North American Quaternary jaguars. In: Martin, R.A., Barnosky, A.D. (Eds.), Morphological Change in Quaternary Mammals in North America. Cambridge University Press, Cambridge, pp. 343-372.
41. Bocherens H. (2015) Isotopic tracking of large carnivore palaeoecology in the mammoth steppe // Quaternary Science Reviews 117, 42-71
p/s. Спасибо Игорю (Corvin) за все замечания
Отредактировано shish02 (23 March 2016 15:27:23)
#275 26 March 2016 23:24:09
- Crazy Zoologist
- Гость
Re: Доисторические пантеры (Pantherini)
Спасибище за такой огромный текст. Обязательно прочту.
Хотел спросить, у пантер во время галопа нет двойного зависания? Нигде про это не читал, а по фильмам судить сложно. Если его нет, то их скорость бега сильно преувеличена. Впрочем. если он есть, но выражен слабо, тоже.