Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#1 28 December 2009 19:48:32
- Crazy Zoologist
- Гость
Пандовые (Ailuridae)
Simocyon.
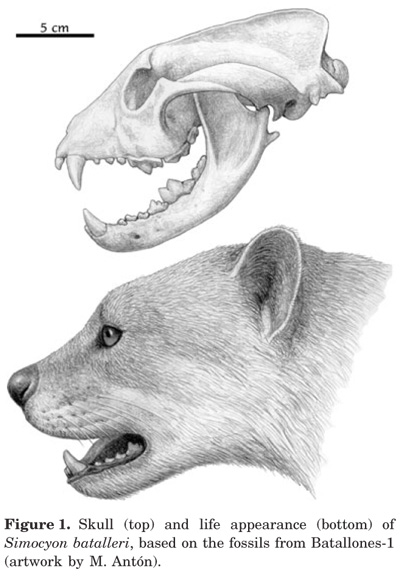
Simocyon, a Unique Extinct Carnivore
THERE ARE certain niches that seem to be filled by one species or another at any time period. In my recent post on mammalian evolution, for instance, I mentioned Castorocauda, a Jurassic mammaliform that seems to have fit into the niche now occupied by beavers or otters. But occasionally we run across an animal that seems to be adapted for a unique role in its ecosystem. One of these animals is Simocyon. This is a puma-sized caniform that lived about 14 million years ago, and died out by four million years ago. Simocyon has a variety of unusual adaptations.
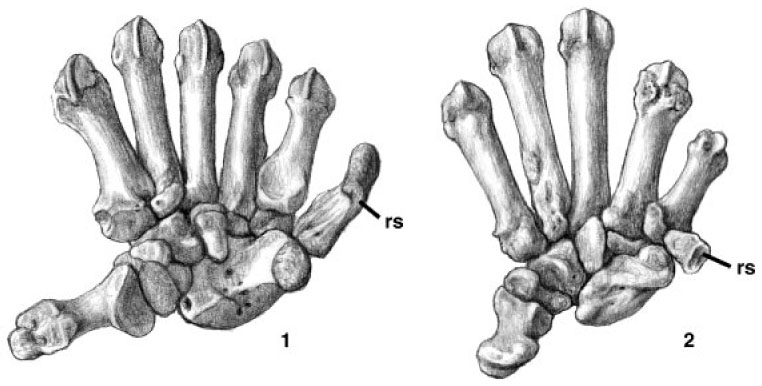
The first unusual feature noted was a false thumb. This character is similar to the thumb of the panda and of the red panda. Since they shared this unusual feature and an odd diet of bamboo it used to be thought that red pandas and pandas are closely related, but it was later discovered that pandas are bears, and red pandas are more closely related to raccoons. The red panda (genus Ailurus) is placed in the family Ailuridae, and Simocyon likewise falls into this family. The false thumbs of the giant panda (1) and Simocyon (2) are shown in the figure to the right.1 The false thumb is produced by an enlarged radial sesamoid bone and serves as a relatively immobile surface that the other digits can be brought into opposition with, allowing a pinching or grasping motion.
In the panda and red panda, the false thumb is a convergently evolved trait and is used to grasp bamboo while eating. Both the panda and red panda have teeth that are adapted for this diet, but Simocyon has more generalized caniform dentition as seen below.2
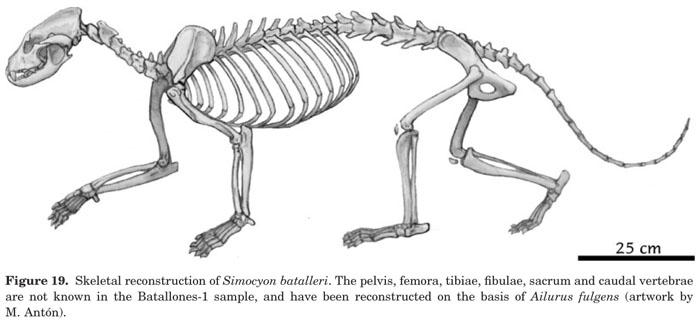
So why the false thumb? It appears that this evolved originally for tree climbing and was preadapted for bamboo eating. Red pandas are primarily arboreal, and Simocyon has skeletal adaptations for climbing. The common ancestor of the two was probably a primarily arboreal carnivore that evolved the false thumb to aid in gripping branches. Salesa and coworkers for the first time have analyzed the postcranial skeleton of Simocyon in detail to determine what its lifestyle was like.2
The odd thing about Simocyon is that it retains many tree-climbing adaptations while being much larger than thought practical for an arboreal carnivore. Indeed, other aspects of its anatomy are more suited to a ground-hunting, cursorial generalized carnivore. The authors suggest that this was Simocyon’s usual mode of behavior, but that it retained its tree-climbing ability because of its unfriendly neighbors. Simocyon lived alongside amphicyonids (colloquially known as bear-dogs) and saber-toothed cats. While Simocyon at the size of a puma was not small (about 54 kg, or 120 lbs), these other carnivores could weigh twice that or more. Simocyon may have scavenged their kills, and its likely these carnivores would have come into contact with Simocyon not infrequently. Large carnivores tend to be aggressive to smaller carnivores, especially those big enough to provide some competition for kills. Lions and hyenas have a mutually antagonistic relationship, wolves kill coyotes, and Great Horned Owls kill smaller owls. So it would not be surprising if most of these confrontations ended up in Simocyon fleeing up a tree. As the authors say, “In this scenario, the strong muscles of the shoulder, forearm and lumbar region of this ailurid would produce the necessary force to propel its body in the vertical faster enough to escape from these encounters.” And I’m sure Simocyon would have been thinking “FASTER!”, so far as it was able to think.
These conclusions were drawn from features of the forelimbs and lumbar spine. The forelimbs possess paws with false thumbs, an increased ability to turn the wrist and hand inward (useful for hugging tree trunks), increased brachiation, and strong muscles in the shoulders. The lumbar spine has unusual adaptations similar to those allowing bounding locomotion in mustelids (think of the way ferrets move), but this type of locomotion is inefficient in an animal this size and this adaptation is probably related to stabilizing the trunk for vertical climbing. At this time the features of the pelvis and hindlimbs are unknown.
It’s interesting to think that the saber-toothed cats may have contributed to the formation of an ecological niche that is absent in modern ecosystems, as well as seemingly occupying one themselves.
(с) http://nimravid.wordpress.com/2008/03/1 … carnivore/
Pristinailurus:
The once mighty red panda empire
In the previous article we looked at the discovery of the Red panda Ailurus fulgens, and also at some aspects of its biology and distribution. There's so much I didn't cover: Red panda physiology is bizarrely interesting, for example. In this article we're going to look at the Red panda's fossil relatives. As I implied in the last article, the Red panda's friends and relations once roamed far and wide. And remember that the term 'panda' belongs to the Red panda and its kin, not to the giant pandas (which are bears, and not close kin of pandas proper). The Red panda and its close relatives belong to a group that we'll be calling the ailurines, and they belong within a more inclusive group which we'll be calling the ailurids...
The first fossil panda to be reported came from... England. In 1888, William Boyd Dawkins described Ailurus anglicus from the Pliocene Red Crag of Suffolk. The Red Crag has a pretty neat fossil mammal fauna: it's also yielded the European puma Puma pardoides, which you'll recall from the European jaguars and pumas article. The jaw fragment described by Dawkins (1888) was about a third bigger in its dimensions than the corresponding part of the A. fulgens jaw, so he described A. anglicus as a 'larger and more powerful animal' than the living species. In 1899, Max Schlosser described a fossil panda skull from Romania, and decided that it was different enough from A. fulgens to deserve its own genus, and the name Parailurus was born. A. anglicus was also referred to this genus. European fossils belonging to Parailurus were later reported from Slovakia and Germany too.
In 1977, Parailurus was reported from North America: Tedford & Gustafson (1977) reported a Parailurus tooth from the Pliocene Ringold Formation of Washington. Sasagawa et al. (2003) later reported Parailurus from the Pliocene Ushigakubi Formation of Japan and Sotnikova (2008) has just described a new species (P. baikalicus) from the Pliocene of Transbaikalia (Russia). Members of this genus seem to have had a wide Holarctic distribution, inhabiting western North America, Asia and Europe at least. Cladistic analysis suggests that Parailurus and Ailurus are sister-taxa (Wallace & Wang 2004), though the possibility that an Asian species of Parailurus gave rise to Ailurus has been suggested at times [adjacent image shows ailurine phylogeny from Wallace & Wang (2004)].
We now know that a second, more primitive ailurine lineage also inhabited North America, thanks to the recent description by Wallace & Wang (2004) of Pristinailurus bristoli from the Upper Miocene/lower Pliocene Gray Fossil Site in Tennessee [life restoration of this species, by Steven Wallace, is shown at the very top]. Wallace & Wang (2004) suggested that this species be known as Bristol's Appalachian panda (the 'Bristol's' refers to Larry Bristol, the finder of the holotype). Originally described from teeth, breaking news for 2008 is that the Gray Fossil site has recently yielded a complete Pristinailurus skull [shown in image below] as well as a reasonable amount of postcranial material. This hasn't yet been published, but you can read about it (and see many images) here. Phylogenetic analysis of tooth characters indicates that Pristinailurus is outside of the ailurine clade that includes Ailurus and Parailurus (Wallace & Wang 2004).
There's no evidence for bamboo at the Gray Fossil Site, raising the question of what Pristinailurus ate. Wallace & Wang (2004) noted that river cane Arundinaria - a member of the bamboo family - is present in Tennessee today, and they suggested that it or a close relative may have been more widespread when Pristinailurus was alive. They also note however that this panda would have had to have gone without bamboo during part of its evolution (during the migration from Eurasia into North America, for example... assuming that that is what it did).
Another fossil ailurine is known from Europe: Magerictis imperialensis, from the Lower Miocene of Spain. It's only known from a single molar unfortunately: this shows that it had a specialized dentition like that of Ailurus and probably had a similar lifestyle. If so, it shows that herbivorous ailurines (and, by inference, their sister-taxon... on which, read on) had evolved not longer after the appearance of the very earliest ailurids in the Late Oligocene.
The simocyonines
So that's the ailurines. Recent discoveries have shown that a long-controversial group of fossil carnivorans, the simocyonines, are also close relatives of Ailurus and should be regarded as part of the same little group, Ailuridae. Originally described in the 1850s and classified alongside a motley assortment of animals now regarded as ursids, canids and mustelids, simocyonines were later identified as part of Canidae, Mustelidae, or Procyonidae, or as relatives of the bear-dogs, or amphicyonids (see Wang 1997 for a review). A list of cranial characters shared with Ailurus allowed Wang (1997) to demonstrate their ailurid identity however. Among these characters are a very strongly arched zygomatic arch, a long external auditory meatus, an erect or anteriorly inclined coronoid process, a particularly big second upper molar, and grooves on the lateral sides of the canines (argh! More mammals with grooved canines!). We currently know of three simocyonine genera (Alopecocyon from Europe, Actiocyon from North America and Simocyon from Europe, Asia and North America), most of which are Middle or Upper Miocene [Chinese Simocyon skull shown in adjacent image].
Unlike ailurines, simocyonines have highly reduced anterior premolars, posterior molars specialized for crushing, and carnassials clearly suited for shearing. These features indicate that, in contrast to ailurines, simocyonines were hypercarnivores and not omnivores or herbivores (Wang 1997, Peigné et al. 2005). Amphictis from the Late Oligocene and Early Miocene of Eurasia, identified by some as an ailurid more basal than both simocyonines and ailurines, also seems to have been hypercarnivorous, so this was always certainly the primitive state for ailurids.
The big crushing posterior premolars of simocyonines have led some authors to suggest that these animals were ecologically equivalent to the bone-crushing borophagine canids (though note that only a few borophagines were like this and, as usual, the group included more diversity than what we're typically exposed to) (e.g. Baskin 1998). Given that simocyonines were clearly good climbers however (read on), they can't have been that much like borophagines. It's possible that they grabbed carcasses or bits of carcasses and then carried them off into trees, but it's also possible that, rather than being crushers of big bones, they used those big posterior premolars to mash up mid-sized prey, bones and all (Peigné et al. 2005).
Long lumbar vertebrae with pointed neural spines and elongate transverse processes show that simocyonines had powerful back muscles which would have provided them with a bounding gait similar to that seen in living mustelids. Puma-sized Simocyon - the best known simocyonine - was surprisingly large for this sort of gait, however, and it's likely that it was built this way because it was a climber rather than a runner. Indeed this is supported by the rest of its skeleton: its shoulders recall those of climbing procyonids like kinkajous (and those of bears, which can also climb), and its forelimbs were flexible and capable of rotation (in cursorial carnivorans, the forelimbs tend to be capable of very little rotation). A new piece of evidence provides further evidence for climbing abilities in simocyonines [adjacent life restoration of Simocyon by Mauricio Antón].
The panda's thumb all over again
Like giant pandas, ailurines possess a 'false thumb': a structure that projects from the wrist (it's actually a radial sesamoid) and is used in gripping the plant stems that the panda then feeds from. The sesamoid was recently described as a rod-like bone lacking a bony articulation with any of the other bones of the hand (Endo et al. 2001), and it has usually been imagined as a specialization for herbivory convergent with that of giant pandas. New data on A. fulgens, however, has demonstrated that the sesamoid is subrectangular (rather than rod-like) with a cartilaginous cap (Antón et al. 2006), and it definitely does have a bony contact with other bones (specifically with the scapholunar).
Furthermore, the discovery of a false thumb in a simocyonine has indicated that this structure did not evolve for use in herbivory given that simocyonines weren't herbivores that wanted to hold on to bamboo stems (so far as we know): Salesa et al. (2006b, 2008) therefore argued that the false thumb evolved early on in carnivorous ailurids as an aid for an arboreal lifestyle, allowing these climbing animals to better grasp small branches, and was later exapted in Ailurus and its close relatives for use in manipulating plant stems. Some simocyonines (like the Spanish species Simocyon battaleri) lived alongside large cats like Paramachairodus and might only have escaped from such animals by retreating to thin branches where the cat was unable to follow (Salesa et al. 2006b, 2008). It has also been suggested that S. battaleri might have eaten from carcasses cached in trees by Paramachairodus [adjacent image shows hand skeletons of Giant panda (on left) compared with that of S. battaleri. 'rs' = radial sesamoid].
Incidentally, we all know that Giant pandas Ailuropoda have 'false thumbs'. But, like Ailurus among the ailurids, we now know that Ailuropoda isn't unique among the ursids in this respect, as the Spectacled bear Tremarctos ornatus also has a very similar large radial sesamoid. Given this phylogenetic distribution it was recently suggested that enlarged radial sesamoids were primitive for ursids but that, while giant pandas and spectacled bears retained them, other ursids reduced them (Salesa et al. 2006a). This discovery (which has been mostly overlooked) is a big deal as it shows that the false thumb of the Giant panda is - like that of the Red panda - apparently inherited from less specialised ancestors, and did not originally evolve for an unusual bamboo-eating lifestyle.
An enduring question about ailurines and simocyonines has been how they're related to other carnivorans, and several different views have been promoted based on different lines of morphological and genetic evidence. I've now written way too much and need to wrap up quickly here, so all I will say is that genetic work on the living Red panda indicates that it and its relative are not procyonids (the group that includes raccoons and coatis), they instead seem to be the most basal branch of the carnivoran clade that includes bears, mustelids and procyonids (Flynn et al. 2000), or the most basal branch of Musteloidea, the clade that includes mustelids and procyonids (Flynn et al. 2005) [adjacent skull reconstruction and life restoration of Simocyon by Mauricio Antón, and from Peigné et al. 2005]
For previous Tet Zoo articles on members of the bear-mustelid-procyonid carnivoran clade (Ursida or Arctoidea or whatever), see the Erongo carcass article, the one on islands of otters and strange foxes, and part II of the Velvet Claw homage. Ailurines and simocyonines have both been in the news in recent years, and quite a few people have written about them. The Voltage Gate covered pandas several times, Carl Zimmer wrote about Simocyon back in 2005, and Nimravid recently wrote about simocyonines here. Roberts & Gittleman (1984) - the classic and most useful reference on the living Red panda - is available for free here, Flynn et al.'s 'Whence the Red panda' can be obtained free here, and Peigné et al.'s 2005 paper on Simocyon can be downloaded for free here.
(с) http://scienceblogs.com/tetrapodzoology … empire.php
Симоцион.

Отредактировано Амфицион (09 July 2017 11:57:35)
#2 28 December 2009 21:39:36

Re: Пандовые (Ailuridae)
Эээ... А есть такое семейство - Малопандовые? Они родственны урсидам? Они ведут своё начало из среднего миоцена или раньше?
- Вы акула пера?
- Нет, я дятел клавиатуры.
Неактивен
#3 28 December 2009 21:42:22
- Arctodus-simus
- Модератор

- Откуда: Лукаморье
- Зарегистрирован: 15 October 2007
- Сообщений: 16362
Re: Пандовые (Ailuridae)
Сейчас это вроде отдельное семейство.
Doctrina multiplex , veritas una !
Неактивен
#4 28 December 2009 23:03:54
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Малые панды - отдельное семейство. Систематика её не вполне ясна. Чисто морфологически она ближе всего к енотам. Некоторые помещают её в одно надсемейство сними, таким образом она ещё дальше удаляется от медвежьих. Другие сближают её с куньими. Согласно молукулярным и биохимическим исследованиям она занимает промежуточное положение между енотами и медведями. Я кстати всегда так и считал. Но согласно другим молекулярным исследованиям она ближе к медведям, согласно третьим - к енотам.
От медведей она отличается строением зубов - бугороков, бакулюмом, ну и прочими чертами, например тем что она полустопоходяща, с длинным хвостом и т.п.
Барышников малых панд делит на три подсемейства: амфиктины, симоционины и собственно малые панды - айлурины.
Род современных малых панд известен из среднего миоцена (Европа). В Сибири - с плиоцена. И что примечательно, в Китае лишь с плейстоцена.
Барышников пишет, что род Pristiailurus очень примитивен, однако он известен (как и пишет Барышников) с позднего миоцена.
#5 29 December 2009 06:40:05
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Семейство малопандовых, или малых панд (Ailuridae) в настоящее время состоит из единственного рода и вида, собственно малой (её ещё называют красной) панды (Ailurus fulgens). Однако в доисторические времена (особенно в миоцене и плиоцене) данное семейство было широко распространено как в Старом, так и Новом Свете и представлено различными родами со множеством видов, весьма разнообразных по внешнему виду и размерам.
Систематика малопандовых в целом и малой панды в частности в своё время вызывала множество научных дискуссий. Её рассматривали в качестве представителя семейства енотовых, Procyonidae (Thenius, 1979; Roberts, Gittleman, 1984), помещали в семейство медвежьих, Ursidae (Hendey, 1980a; Mayr, 1986; Wozencraft, 1989a, 1993; Wyss, Flynn, 1993) или же выделяли в отдельное семейство малопандовых, Ailuridae (Pocock, 1941; Baskin, 1982, 1989; Grinsburg, 1982).
Дальше тут: http://crazy-zoologist.livejournal.com/463214.html
#6 29 December 2009 14:32:13
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
То же что написано в моей статье (по Барышникову).
#7 19 December 2010 18:23:16
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Никому не известны другие представители семейства?
Интересно получается, что в древности малопандовые были настоящими хищниками. Череп симоциона - 100% принадлежит активному хищнику. Размером он был с пуму. Потом по каким-то причинам они перешли на растительную диету.
#8 20 December 2010 13:45:21
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
В википедии пишут что на симоциона охотились нимравиды, ранние псовые итремаркторины. Но не каждый нимравид был крупнее симоциона, который был размером с пуму. К тому же это был настоящий хищник.
У него был ещё карликовый родственник:
Alopecocyon was a small relative of the modern Red Panda. It weighed only about 11 lbs (5 kg). It was closely related to Simocyon, a larger member of its group. Its fossils have been found in France, Poland, and Slovakia.
#9 05 February 2011 22:28:23
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Не позже. А какой самый ранний не знаю.
#10 23 July 2011 15:24:25
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
А давно малые панды в мустелоидах?
#11 12 April 2012 19:44:50
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Пандовые (Ailuridae)
Оживляем тему. Перевод статьи Матишова и Калмыкова «Red Pandas (Mammalia, Carnivora: Parailurus) in the Biomes of North Eurasia and North America», 2011 (не могли они сразу на родном написать – русскоязычных авторов с америкосовского переводим, нонсенс!)
Находка ископаемых остатков красной панды (Parailurus) на западе Забайкалья имеет особое значение по сравнению с другими открытиями азиатских енотов (Ailurinae), поскольку она позволяет соединить биомы Европы, Азии и Северной Америки, существовавшие в течение позднего неогена. Нынешний ареал обитания Ailurus fulgens ограничен Юньнанем и Сычуанем в Китае, Северной Мьянме, Бутане, Непале и Северо-Восточной Индии, где она обитает в высокогорных широколиственных и хвойных лесах умеренного климата с кустами бамбука и рододендрона.
Один из авторов обнаружил фрагменты красной панды на южных склонах хребта Кхамба, Западного Забайкалья, где млекопитающие из в предыдущие эпохи обитали млекопитающие из отрядов зайцеобразных, грызунов, приматов, хищных, хоботных, непарно- и парнокопытных жили. Это открытие представляет интерес не только с точки зрения изучения таксономической принадлежности и появления плиоценовых азиатских енотов в Евразии, а также среды их обитания.
Особенности рельефа, климата и растительности в раннем плиоцене демонстрируют, что условия, в которых млекопитающие обитали в Западном Забайкалье (Удунг) являлись уникальными в течение позднего плиоцена и в более поздние периоды в Центральной Азии. В плиоцене горная область Западного Забайкалья и Северной Монголии характеризуется вертикальной зональностью растительного покрова, о чём свидетельствует появление не только хвойных (тсуга, ель, сосна и лиственница), но и лиственных (вяз, липа, дуб, граб, орех, ольха, лещина, хмель, береза и ива) лесов. Нижний уровень в этих местах был представлена сфагновыми, плауновыми, папоротниковыми и смородинными. Это подтверждается открытиями представителей уникальной фауны, представленной церкопитеками, пандой, куницей, росомахой,медведем, пахикрокутой, гомотерием, мамонтом, южным слоном, схизотерями, гиппарионом, носорогом, оленями - аксисом и орхоноцеросом, антилопами, газелями, баранами и другими родами.

Ископаемые остатки красных панд: фрагмент верхней челюсти (а, б) и нижней (г) Parailurus baikalicus, Россия. (с) нижняя челюсть Pristinailurus bristoli, США.
В Европе, бамбук ("Bambusa" lugdunensis) был широко распространён в начале палеогена в восточной части Балканского полуострова. В период позднего тортония, Bambusa iljinskia появился в районе Прикарпатья, где он (вместе с рододендроном и корицей) входил в состав флоры, которая состояла из плаунков, плауновидных, древовидных и наземныхпапоротников, пихты, тсуги, ели, кедра, сосны, восковницы, ореха, лещины, берёзы, ольхи, бука, каштана, липы и клёна. В сармате "Bambusa" lugdunensis являлся обычным элементом флоры Альп и на Балканах, а в течение плиоцена в горных группах Альп, Апеннин, Закарпатской области и также северо-западной части Закавказья. Представители другого рода бамбука (Sasa kodorica) росли не только в олигоцене в восточной части Балканского полуострова и в плиоцене в северо-западной части Закавказья и закарпатской области - в настоящее время он широко распространён на Курильских островах и Южном Сахалине. Бамбук является неотъемлемой частью леса в южной части Центральной и Западной Европы (Альпы) и юге Европы (Апеннины) в период с олигоцена до среднего плейстоцена и в Восточной Европе (Карпаты), в течение неогена. В среднем и позднем миоцене бамбук был довольно широко распространен в Восточной Европе (Судеты и Карпаты), что подтверждается многочисленными открытиями лиственных отпечатков и пыльцы "Bambusa" lugdunensis и Graminidites bambusoides. В позднем миоцене флора Восточной Европы (Карпаты) состояла не только из мхов, папоротников, водорослей и травянистых растений, но и хвойных (пихта, сосна, кедр, кетелеерия, ель, секвойя, сциадопитис и тсуга) и широколиственных (клён, ольха, граб, орех, каркас, бук, липа, ликвидамбар, птерокария, луб, вяз, березы и ивы) деревьев. Нижний уровень в лесах был представлен рододендроном (Ericipites roboreus) и бамбуком (Graminidites bambusoides). В плиоцене, кроме Bambusa lugdunensis флора Закарпатья включала роды и виды, характерные для тургайской флоры. Аналогичная флора была широко распространена практически во всей Голарктике, от Британских островов до Японии, где были обнаружены фрагментарные находки плиоценового рода красной панды Parailurus.
Среда обитания современной Ailurus fulgens, её пищевые адаптации, а также тип зубов не имеют принципиальных отличий от зубов плиоценовых представителей родов Parailurus и Pristinailurus bristoli в Северной Америке в эпоху позднего миоцена. Ряд других факторов также доказывает, что представители Parailurus (anglicus, P. hungaricus и P. baikalicus) жили в одинаковых условиях. В большинстве работ, посвящённых систематике и генеалогии рода Parailurus Европы, Азии и Северной Америки, основным направлением является описание морфологических особенностей отдельных Р4 и М1, который раскрывают особенности их индивидуальной, географической и гомологической изменчивости, адаптаций питания и среды обитания. Пищевые адаптации древней красной панды, как и современной, в первую очередь связаны с бамбуковой диетой, несмотря на некоторые усилия, это опровергающие. Когда красные панды жили в Европе, в тамошних лесах произрастал бамбук и кусты рододендрона - современная A. fulgens тоже предпочитает такие места.
Раннеплиоценовая флора Хамбо-Ридж имеет ряд общих таксонов с европейской флорой неогена (сфагновые, плауновые, тсуга, ель, сосна,вяз, липа, дуб, клён, ольха, граб, лещина, орех и ива), но не включает бамбук или реликты полтавской флоры, которые являются типичными для европейской флоры в неогене. Отсутствие пыльцы бамбука и отпечатков листьев не только в Забайкалье, но и в других местах, где были обнаружены фрагментарные остатки красной панды, можно объяснить небольшим количеством образцов, взятых для анализа и литологической характеристикой отложений, что помешало сохраниться хрупкой пыльце бамбука. В Японии, где бамбук растет в настоящее время, красные панды (Parailurus sр.) были обнаружены в отложениях плиоцена в префектуре Ниигата. Они жили на островах 4-3 млн лет назад и имели общие черты P4 как у современных A. fulgens и плиоценовых представителей Parailurus, которые жили на континенте почти в то же время.
В другой части Голарктики, в Северной Америке, красные панды были обнаружены в верхнем миоцене до отложений нижнего плиоцена в штате Теннесси (Pristinailurus bristoli) и в отложениях верхнего плиоцена в штате Вашингтон (Parailurus sр. ). Леса были похожи на те, где были широко распространены различные субтропические млекопитающие (Arctomeles, Machairodus, Plionarctos, Gomphotherium и Tapiravus, Teleoceras) в восточной части Соединенных Штатов 7.0-4.5 млн лет назад. Таким образом, открытие красной панды (P. baikalicus) на западе Забайкалья доказывает, что тургайская флора была широко распространена в эпоху позднего неогена, что позволило многим лесным млекопитающим, включая красную панду, распространиться почти на всю Голарктику. Пространственный разрыв обитания ископаемой красной панды не является отражением только топографических особенностей скудность палеонтологической летописи, - это также отражает и небольшие размеры её популяций. Неуклонное похолодание климата, прогрессивное сглаживание рельефа и другие факторы деструктивно повлияли на ареал их обитания в течение позднего плиоцена и красные панды начали вымирать, сохранившись только в Китае.
http://www.springerlink.com/content/qk37333jh37kj24n/
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#12 13 April 2012 16:09:18
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Пандовые (Ailuridae)
Кот :
А давно малые панды в мустелоидах?
Айлуриды включены в надсемейство Musteloidea - каниформ, куда помимо Ailuridae (малопандовых) входят Mephitidae (скунсы), Mustelidae (куньи) и Procyonidae (енотовые) Флинном в 2001 г. Относительно внятные объяснения я раньше только у Гинсбурга читал, но сейчас под рукой есть масса инфы, проливающей свет. Надеюсь, не будет возражений, если я в нескольких постах (для визуального удобства восприятия, чтобы не загромождать одним объёмным) изложу филогению айлурид.
Классификация красной панды стала предметом многочисленных дискуссий с самого начала первоначального описания Кювье (1825). Морфологические и молекулярные исследования подтвердили широкий спектр возможных отношений, в том числе тесные связи с Ursidae, Procyonidae и Mephitidae. Тем не менее, другие исследования доказывают, что красная панда может лежать в основе клад, включающих Procyonidae+Mephitidae+Mustelidae; Ursidae+Procyonidae; Ursidae+Otariidae или Ursidae+Pinnipedia. В результате этих исследований красная панда классифицировалась как урсид, проционид или же в рамках собственного семейства, Ailuridae. При рассмотрении вопроса о филогении все усилия сосредотачивались на остеологии красной панды и краниодентальных элементов в частности. Функции мягких тканей в использовании филогенетического анализа и миологии изучались мало.
Изначально Ailurus связывали с Procyonidae. Мак-Гру (1938) описал сходство молярной модели цинарктоидов, первых проционид миоцена и плиоцена Северной Америки с Ailurus. Phlaocyon (нижний-средний миоцен Северной Америки) и Aletocyon (нижний миоцен Северной Америки) имеют существенные сходства с Ailurus на основе моляров, хищнических зубов и структуры слуховой области. Симпсон (1945) предположил, что Phlaocyon обладал несколько промежуточными особенностями между Cynodictis и Procyon и применил те же аргументы в отношении Aletocyon, разместив оба рода в подсемейство Procyoninae и сделал вывод, что они скорее являлись предшественниками общей радиации проционид, чем айлурид.
По современным представлениям, самые ранние предки красных панд были миациды из позднего палеоцена и эоцена Евразии и Северной Америки, примерно 55-34 миллионов лет назад – парафилетическая группа, хорошо приспособленная к обитанию в пышных эоценовых лесах. В конце эоцена они привели к двум отдельным группам - геспероционинам, ранним псовым с одной стороны, и к урсидам в лице американского рода Mustelavus, от которого, предположительно, также дихотомировали айлуриды и проциониды. Таким образом в дальнейшем структура выглядела так:
Семейство Ailuridae
-(†)Род Amphictis
-(†)Род Protursus
(†)Подсемейство Simocyoninae
Подсемейство Ailurinae

Собственно айлуриды, вероятно, появились в Европе в позднем миоцене – раннем олигоцене (примерно 25-18 млн лет) - род Amphictis Pomel, 1853. Животное с черепом длиной 10 см, вероятно, соразмерное современной красной панде, хотя относительно хорошо известны только черепа и зубы. Всеядный хищник с простыми премолярами, относительно острыми гранями хищнической пары (Р4 и М1) и перетирающей поверхностью моляров. Хотя во многих отношениях (расположение зубов, базикраниум) морфологии Amphictis остается примитивным, удлинение m2, который также имеет полностью разделённые протоконид и метаконид и наличествуют зачаточные боковые ложбинки на клыках, поддерживает размещение Amphictis в Ailuridae. Очевидно, его диета состояла из мелких позвоночных, беспозвоночных, яиц и фруктов. Даже если отношения Amphictis к линии красной панды, айлурин, ещё требует дополнительных подтверждений, то с симоционинами, особенно с Alopecocyon филогенетически они очень близки.

Череп Amphictis
Отредактировано Сorvin (13 April 2012 16:11:34)
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#13 13 April 2012 16:24:32
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Пандовые (Ailuridae)
(†)Подсемейство Simocyoninae
-(†)Род Alopecocyon (Actiocyon?)
-(†)Род Simocyon
Alopecocyon известен по фрагментарному материалу из среднего миоцена Европы, Азии и Северной Америки (под названием Actiocyon – вероятно, младший синоним). Alopecocyon отличается от Amphictis в том, вторые моляры расширены по сравнению с первыми коренными зубами, обладает иной структурой m2 и более глубокими боковыми пазами на клыках. Тесная связь между Alopecocyon и Simocyo признана уже давно, основываясь на базикраниальном материале, дентальных характеристиках и посткраниали.
Simocyon, размером с пуму - один из хорошо известных ископаемых айлурид, известный из среднего и позднего миоцена Северной Америки и Европы, а также из среднего миоцена - раннего плиоцена Китая. Его краниодентальная анатомия хорошо известно благодаря полным черепам из Китая и Испании, ясно размещающая Simocyon в семейство Ailuridae и позволяющая проследить в этом роде дентальную эволюцию. Более примитивный вид Simocyon batalleri валлезия (поздний миоцен, 11,1 - 8,7 миллионов лет назад) обладал полным набором зубов, относительно небольшими клыками, четырьмя премолярами и двумя коренными зубами и в верхней и нижней челюсти, в то время как у более позднего вида Simocyon primigenius из туролия (поздний миоцен, 8,7 - 5,3 миллионов лет назад) наблюдается редукция числа премоляров, но с тем же числом коренных зубов, как и у S. batalleri. Скорее всего, это сокращение не связано с адаптацией к травоядной пище наряду с ярко выраженными хищническими зубами. Таким образом, Simocyon, вероятно – всеядный хищник, который охотился на мелких позвоночных, но мог также потреблять фрукты, семена и яйца.
A - Simocyon cf. batalleri (рисунок М. Антона), B, C - Simocyon primigenius (реконструкция В. Симеоновски)
В частности, большинство посткраниальных специализаций, присутствующих у S. batalleri предполагают неплохие способности к древолазанию, хотя, учитывая предполагаемые размеры этого животного около 60 кг, трудно представить его древесным жителем типа красной панды. Но одной из самых поразительных особенностей, которой обладал S. batalleri, является так называемый "ложный большой палец", гипертрофированная кость запястья (радиальная сесамовидная), отчасти напоминающая функционально отстоящий большой палец. Эта черта также присутствует как у гигантской панды (Ailuropoda melanoleuca), так и у красной панды, использующих эту структуру для сдирания листьев с бамбуковых ветвей, составляющих основную часть их рациона. Ранее эта особенность использовалась для предположения о тесной взаимосвязи современных панд. Тем не менее, последние молекулярные исследования показали, что гигантская панда является медведем. Другими словами, эта специализированная структура имела различное происхождение в этих видах и на самом деле изучение функциональной морфологии показывает ясные отличия в форме, относительных размерах и даже мышечных приложений.
Более вероятно, что это животное использовало свои способности к древолазанию как способ уйти от крупных хищников позднего миоцена - амфиционид или саблезубых кошек. Агрессивные по отношению к потенциальным конкурентам они могли быть очень опасны для таких среднеразмерных и относительно уязвимых животных, как S. batalleri. Анатомия этого животного указывает на наличие сильных мышц плеча, предплечья и поясничной области, продвигающих тело вертикально достаточно быстро, чтобы избавиться от этих встреч. В этом контексте «ложные пальцы" приобретают функции своего рода клещей, позволяющих S. batalleri достичь относительно тонких ветвей, где он может спастись от крупных хищников, а также, в свою очередь, охотиться на мелких позвоночных - ящериц, птиц и грызунов.
Скелетная анатомия Simocyon batalleri: (A) реконструкция S.batalleri. (B) Запястные и пястные кости, показывающие положение "ложного пальца" (РС) у гигантской панды (Ailuropoda melanoleuca) (слева, 1) и S. batalleri (справа, 2) (без соблюдения масштабов).
Отредактировано Сorvin (13 April 2012 16:29:02)
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#14 13 April 2012 16:26:41
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Пандовые (Ailuridae)
Подсемейство Ailurinae
-Род Ailurus
-(†)Род Magerictis
-(†)Род Parailurus
-(†)Род Pristinailurus
Ранние айлуриды (Amphictis и Alopecocyon) не обладают специализированной дентальной морфологией, свидетельствующей о растительноядности, присутствующей у представителей подсемейства Ailurinae: Pristinailurus, Parailurus и Ailurus. Другой айлурид, наряду с Alopecocyon очень близкий к Amphictis - миоценовый Magerictis imperialensis. Этот вид был описан на основе отдельного m2, обнаруженного в Испании. Морфология этого зуба, ясно принадлежащая айлуридам, демонстрирует очень удлинённую коронку и более чёткое разделение метаконида и протоконида (более ясно, чем у Amphictis) по продольному углублению, которое продолжается по всей длине зуба.
Представители Ailurinae адаптированы к диете, состоящей в основном, из растительных материалов и обладают совершенно другим дентальным шаблоном - окклюзионными поверхностями из нескольких затуплённых бугорков, что очень похоже на особенности, имеющиеся больше у свинообразных, чем у хищников. Эта параболическая модель позволяет эффективно измельчать пищу и очень эффективна при обработке такой жёсткой растительности, как листья бамбука, составляющих основной рацион современных красных панд.
Несмотря на распространение современных Ailurus лесами умеренного пояса горного южного региона Гималаев, ископаемые формы подсемейства были широко распространены в Европе в Восточной Евразии. Magerictis imperialensis существовал в течение среднего миоцена Испании; Parailurus angliеus в плиоцене Англии, Румынии, Словакии и Германии; Parailurus hungaricus известен из плиоцена Словакии, Parailurus baikalicus обнаружен в среднеплиоценовых отложениях Забайкалья. Два изолированных верхних зуба Parailurus sр. были обнаружены в плиоценовых слоях Северной Америки и Японии. Pristinailurus bristoli, с конце миоцена до раннего плиоцена, был обнаружен в Северной Америке.
Карта, показывающая распределение Ailurinae. Местоположения: Parailurus – красный цвет, Pristinailurus bristoli – синий, Magerictis – жёлтый. Современное распределение Ailurus - зелёный цвет с двумя подвидами: А. f. fulgens – тёмно-зелёный и А. f. styani – светло-зелёный.
Промежуточные формы между Parailurus и Ailurus не известны. Имеющиеся материалы по Parailurus свидетельствуют, что все три разных вида были крупнее и массивнее, чем Ailurus. Тедфорд и Густафсон (1977) предполагают, что североамериканский Parailurus, вероятно, распространился из Азии. Его находки в Японии подтверждают гипотезу о том, что Parailurus в плиоцене был широко распространён во всём северном умеренном поясе.
Палинологические данные свидетельствуют о том, что климат, в котором жил Parailurus, был прохладный и умеренный. Вполне возможно, что [i]Ailurus произошёл от какой-то популяции широко распространённого Parailurus (Гинзбург и др., 1997) и остались ограниченным ареалом в субтропических высокогорьях Азии. Другие считают Parailurus слишком прогрессивными, чтобы служить предками Ailurus (например, Ванг, 1997). Меньший размер и уменьшение ареала Ailurus также может говорить о том, что красная панда может представлять собой специализированное ответвление линии айлурин (и, возможно, даже азиатскую форму Parailurus), которое пережило плейстоценовые оледенения в горных убежищах на юге Китая.
Приватный вывод – красная панда сравнительно произвольная форма, довольно далёкая от морфотипичных представителей, если учитывать краниальные и дентальные адаптации для растительного рациона. Но, несмотря на редкие и фрагментарные находки, общая картина более-менее ясна. К сожалению, почти ничего не знаю по базальному айлуриду Protursus. И, краем глаза, встречал информацию, что Sivanasua из верхнего миоцена Европы и нижнего плиоцена Азии, возможно, также является примитивным представителем подсемейства Ailurinae.
ЛИТЕРАТУРА:
-Anton M. et al.. Implications of the functional anatomy of the hand and forearm of Ailurus fulgens (Carnivora, Ailuridae) for the evolution of the ‘false-thumb’ in pandas. 2006
-Fisher R. et al. The phylogeny of the red panda (Ailurus fulgens) evidence from the hindlimb. 2008
-Ginsburg L. et al. Un Ailurinae (Mammalia, Carnivora, Ailuridae) dans le Miocene moyen de Four (Isere, France). 2001
-Glatston A. Red panda - biology and conservation of the first panda. 2011
-Matishov. Kalmykov P. Red Pandas (Mammalia, Carnivora Parailurus) in the Biomes of North Eurasia and North America. 2011
-Ogino S. et al. Mandible and Lower Dentition of Parailurus baikalicus (Ailuridae, Carnivora) from Transbaikal area, Russia. 2009
-Peigne S. et al. Ailurid carnivoran mammal Simocyon from the Late Miocene of Spain and the systematics of the genus. 2005
-Roberts M., Gittleman J. Ailurus fulgens.1984
Отредактировано Сorvin (13 April 2012 16:30:06)
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#15 14 April 2012 00:59:02
- Юра
- Без пяти минут зоолог
- Зарегистрирован: 31 July 2008
- Сообщений: 10735
Re: Пандовые (Ailuridae)
Неактивен
#16 13 July 2012 02:03:03
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Корвин, получается, что еноты ближе к хорьковым, чем к медвежьим?
#18 06 June 2017 23:28:07
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Семейство малопандовых, или малых панд (Ailuridae) в настоящее время состоит из единственного рода и вида, собственно малой (её ещё называют красной) панды (Ailurus fulgens). Однако в доисторические времена (особенно в миоцене и плиоцене) данное семейство было широко распространено как в Старом, так и Новом Свете и представлено различными родами со множеством видов, весьма разнообразных по внешнему виду и размерам.
Систематика малопандовых в целом и малой панды в частности в своё время вызывала множество научных дискуссий. Её рассматривали в качестве представителя семейства енотовых, Procyonidae (Thenius, 1979; Roberts, Gittleman, 1984), помещали в семейство медвежьих, Ursidae (Hendey, 1980a; Mayr, 1986; Wozencraft, 1989a, 1993; Wyss, Flynn, 1993) или же выделяли в отдельное семейство малопандовых, Ailuridae (Pocock, 1941; Baskin, 1982, 1989; Grinsburg, 1982).
Биохимические и молекулярные исследования не однозначно определяют систематическое положение данного вида. Согласно O'Brien et al. (1985), Tagle et al. (1986) и Wayne et al. (1989) малая панда занимает промежуточное положение между енотовыми и медвежьими. Согласно Braunitzer (1987), Hofmann (1987), Goldman et al. (1989), Slattery (1995), O'Brian (1995), Lan и Wang (1998) она ближе к енотовым, а по мнению Zhang (1991), Shi (1991) и Vrana et al. (1994) ближе к медвежьим. Более новые данные молекулярной биологии сблидают малую панду со скунсовыми (Mephitidae) и помещают в сестринский таксон к семейства куньих, Mustelidae, и енотовых, Procyonidae (Delisle, Strobeck, 2005; Flynn et al., 2005). Таким образом, согласно современной систематике, малую панду выделяют в отдельное семейство Ailuridae и надсемейство Musteloidea (куницеобразные, или куницеподобные), сближая их тем самым со скунсовыми (Mephitidae), куньими (Mustelidae) и енотовыми (Procyonidae).
Морфологически малая панда больше всего похожа на енота, однако у неё отстутвует большинство производных признаков, типичных для последних. Некоторые анатомические черты роднят её с представителями семейства медвежьих, например, строение слухового пузыря, но у малой панды присуствуют многие черты, не свойственные медведям - длинные вибриссы, хорошо развитые анальные железы, иные пропорции конечностей, слабое развитие плантарных подушечек, полувтяжные когти, иное строение бакулюма, длинный хвост, слабо развитые губы, иная зубная морфология и т.п.
Семейство малопандовых делят на три подсемейства - Ailurinae, Simocyoninae и Amphictinae (Grinsburg et al., 1997, 2001; Baskin, 1998).
Возраст ископаемых находок представителей подсемейства Ailurinae, таких как Magerictis и Parailurus ограничен средним миоценом Европы (Grinsburg et al., 2001). Саммые ранние останки рода Ailurus (к которму принадлежит современная малая панда) относятся к плиоцену Сибири (Калмыков, 1999). Примечательно, что в Китае Ailurus появляется лишь в плейстоцене (Bien, Chia, 1936). Из Северной Америки известны очень примитивный Pristinailurus из позднего миоцена или раннего плиоцена штата Тенесси (Wallace, Wang, 2004) и Parailurus, известный по единственному верхнему моляру из позднеплиоценовых отложений штата Вашингтон (Tedford, Gustafson, 1977).
Из этого можно заключить, что семейство малых панд в прошлом имело преимущественно голарктическое распространение, а их современный ареал, ограниченный небольшими районами горных лесов Южной Азии является реликтовым.
#19 09 June 2017 14:07:14
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Ailurid carnivoran mammal Simocyon from the late Miocene of Spain and the systematics of the genus
https://www.app.pan.pl/archive/publishe … 50-219.pdf
#20 07 July 2017 19:23:13
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
А что значит "радиальная сесамовидная"? Радиальный - обращенный к радиусу. Она что, к радлиусу обращена? Че-то я туплю, зхвиняйте и объясните если не лень.
А говорят, что он был гиперхищником, но судя по нижним молярам он явно ел и растительную. пищу тоже. Что скажете?
Отредактировано Амфицион (07 July 2017 19:24:23)
#21 22 September 2017 19:56:15
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Шестой палец запястья это именно радиальная сесамовидная кость?
Кстати у малой панды подошвы лап покрыты густыми волосами. Это весьма странно для древолазающего животного.
Отредактировано Амфицион (22 September 2017 20:23:11)
#22 22 September 2017 20:48:37
- Sitta europaea
- Без пяти минут зоолог

- Зарегистрирован: 07 February 2016
- Сообщений: 1818
Re: Пандовые (Ailuridae)
Амфицион :
А что значит "радиальная сесамовидная"? Радиальный - обращенный к радиусу. Она что, к радлиусу обращена? Че-то я туплю, зхвиняйте и объясните если не лень.
https://upload.wikimedia.org/wikipedia/ … genius.JPG
А говорят, что он был гиперхищником, но судя по нижним молярам он явно ел и растительную. пищу тоже. Что скажете?
Точно не хищнее лисы.
Вожаки серых волков воют в ивняке,
Опавшие листья летят, уносимые ветром.
Неактивен
#23 22 September 2017 21:36:43
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Red pandas live in temperate climates in deciduous and coniferous forests. There is usually an understory of bamboo and hollow trees. The average temperature is 10 to 25 degrees Celsius, and the average annual rainfall is 350 centimeters. (Glatston 1994, Roberts and Gittleman 1984)
Как? Если она обитает на высоте от 2200 до 4800 метров? К тому же в этой же статье пишется что обитает она в умеренном климате. Данные из ADW.
#24 23 September 2017 17:47:25
- Crazy Zoologist
- Гость
Re: Пандовые (Ailuridae)
Малая, или красная панда (Ailurus fulgens) представляет собой живое ископаемое, поскольку это единственный доживший до наших дней вид семейства пандовых (Ailuridae), которые были более-менее широко распространены миллионы лет назад. Малая панда находится лишь в отдаленном родстве с большой пандой (Ailuropoda melanoleuca), поскольку она, естественно, более тесно связана с другими членами надсемейства Musteloidea, к которому она принадлежит, в отличие от большой панды, которая относится к надсемейству Ursoidea.
Общим предком обеих панд (который также был предком для медведей, ластоногих, а также членов надсемейства Musteloidea, к которому принадлежат куньи, скунсовые, енотовые и пандовые) обитал в раннем третичном периоде десятки миллионов лет назад и имел широкое распространение по всей Евразии.
Окаменелости ископаемой красной панды Parailurus anglicus были обнаружены на востоке в Китае и на западе в Великобритании. В 1977 году в плиоценовых отложениях Ringold Formation в Вашингтоне был обнаружен единственный зуб Parailurus. Эта находка почти идентична европейским образцам и указывает на иммиграцию этого вида/рода из Азии в Северную Америку. Это означает, что представители рода Parailurus имели очень широкое распространение в плиоцене. Кладистический анализ показывает, что Parailurus и Ailurus являются сестринскими таксонами.
В 2004 году в Gray Fossil Site в Теннесси был обнаружен зуб, принадлежащий одному из видов красных панд, ранее не зарегистрированный в Северной Америке. Данный зуб датируется 4,5-7 миллионов лет назад (я не понимаю, откуда такая разница в датировен). Этот вид, описанный как Pristinailurus bristoli, указывает на то, что вторая, более примитивная линия Ailurinae населяла Северную Америку во время миоцена.
Дополнительные окаменелости Pristinailurus bristoli были обнаружены в Gray Fossil Site в 2010 и 2012 годах. Частота находок окаменелости этой панды в Gray Fossil Site предполагает, что данный вид играл важную роль в экосистеме этого района.
Открытие в Испании посткраниальных останков Simocyon batalleri (миоценового родственника малой панды из другого подсемейства) подтверждают сестринские отношения между пандовыми и медвежьими. Данное открытие говорит о том, что ложный палец развился у пандовых и большой панды независимо. Большой панде он нужен для удержания тонких побегов бамбука, а рпндовым для древолазания (с этим я не совсем согласен, так как современная малая панда также как и большая использует его, помимо лазания по деревьям, для удержания ветвей бамбука).