Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#951 30 December 2018 13:07:41
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Подсемейство BOROPHAGINAE Simpson, 1945
РАСПРОСТРАНЕНИЕ: ореллий – бланк Северной Америки до хемфиллия Центральной.
ДИАГНОЗ: борофагины и канины отличаются от геспероционов в дальнейшем уменьшении парастиля на M1, удлинённого переднего внутреннего цингулума на M1, редуцированного талонида на m1 и метаконида на m2, сближенного с протоконидом. По сравнению с канинами у борофагов имеются определённые синапоморфии: удлинённая мандибула, узкие и длинные разделённые диастемами премоляры, редуцированные или отсутствующие задние дополнительные вершинки на премолярах, лонгированный тригонид на m1 и удлинённые m2. Хотя не исключено, что большинство из этих символов, очевидно, появились у борофагин в процессе эволюции адаптивно и независимо.
ARCHAEOCYON Wang,1999
РАСПРОСТРАНЕНИЕ: поздний уитний - средний арикарий Северной Америки, 30-25 млн лет назад (ранний олигоцен-ранний миоцен).
ДИАГНОЗ: в качестве базального таксона Archaeocyon демонстрирует все синапоморфии, которые объединяют подсемейства Caninae/Borophaginae и дифференцируют их от геспероционин: небольшой парастиль на M1, окружённый протоконом лингвальный цингулум на M1 спереди, базальный талонид на m1 и метаконид на m2 немного выше, чем протоконид. В отличие от Caninae Archaeocyon не имеет производных символов для этой группы и остаётся примитивным по отношению к большинству представителей борофагин.
ОБСУЖДЕНИЕ: по-видимому, Archaeocyon является базальным таксоном как для Borophaginae, так и для Caninae. И, подобно Hesperocyon, также является парафилетическим родом, не проявляющим явной морфологической тенденции к более прогрессивным линиям. Учитывая, что зубная морфология базальных борофагов имеет тенденцию быть более консервативной (т. е. более близкой к Hesperocyon), чем их аналог в Caninae (Leptocyon), Уонг (Wang, 1999) размещает Archaeocyon в борофагинах вместе с Oxetocyon и Otarocyon, которые занимают аналогичную двусмысленную позицию в кладограмме.
Это логично, потому что самое раннее появление Leptocyon фиксируется в ореллании (ранний олигоцен) - таким образом, маловероятно, что Archaeocyon является явным предком Caninae. Немногие прогрессивные дентальные признаки археоциона (узкий талонид, расширенный лингвальный цигулум на М1 и т. д.) направлены на первоначальную тенденцию к гиперкарниворности относительно такового состояния в Hesperocyon. Подобная тенденция позволила впоследствии борофагам занять более высокие трофические ниши в сравнении с геспероционинами.
Archaeocyon pavidus
ГОЛОТИП: LACM-CIT 466, раздробленная передняя половина черепа и нижняя челюсть из Калифорнии.
РАСПРОСТРАНЕНИЕ: поздний уитний - ранний арикарий Северной Америки.
ОБСУЖДЕНИЕ: помимо некоторых дентальных особенностей, которые объединяют его с борофагинами, A. pavidus скорее схож с Hesperocyon по размеру и пропорциям. A. pavidus был признан, возможно, как очень примитивным борофагином. Уонг и Тедфорд (Wang and Tedford, 1996) даже временно именовали его как Cormocyon pavidus, отмечая базовую «барофагинность».
Archaeocyon leptodus 
ГОЛОТИП: MCZ 2878 - левый частичный череп со сравнительно полным зубным рядом
РАСПРОСТРАНЕНИЕ: ранний арикарий Северной Америки.
ОБСУЖДЕНИЕ: образцы A. leptodus трудно отличаются от уитнийской формы как по морфологии, так и по метрике. Основное отличие заключается в нескольких нюансах: более заднеориентированный парокципитальный выступ, чуть более крупная булла, гипокон на M1 позиционируется более постериорно, на M2 нарушена связь между метаконулем и внутренним цингулумом, m1 более удлинённый. Таким образом, наличествует две сходных, но морфологически различимых формы.
Archaeocyon falkenbachi 
Archaeocyon falkenbachi (FAM 49029), голотип
ГОЛОТИП: FAM 49029, фрагментарный череп с частичным зубным рядом.
РАСПРОСТРАНЕНИЕ: средний арикарий Северной Америки.
ОБСУЖДЕНИЕ: небольшой износ на вершинах верхних коренных зубов и наличие низкого, но отчётливого сагиттального гребня свидетельствует, что образец принадлежит половозрелой особи и маловероятно, что его брахицефальные краниальные пропорции обусловлены ювенильностью. Таким образом, A. falkenbachi представляет собой отдельную филогенетическую линию, отличающуюся от A. leptodus.
OXETOCYON Green, 1954
Oxetocyon cuspidatus (SDSM 2980), голотип
ДИАГНОЗ: в отличие от Archecheocyon, Oxetocyon разделяет с Otarоcyon такие производные характеристики, как более высокий парастиль на P4, раздельные энтоконид и метаконид на m1 и метаконид больше, чем протоконид. Кроме того, Oxetocyon обладает несколькими аутапоморфиями, связанными с его специфической адаптивностью к гиперкарниворности: увеличенные и квадратные верхние моляры, массивный передний цингулум на M1 и выраженный метаконуль на M1. Oxetocyon примитивен по отношению к Rhizocyon и более прогрессивным борофагинам в отсутствии связи между метаконулем и задним лингвальным цингулумом на M2, а также отсутствием протостилида на m1.
Oxetocyon cuspidatus
ГОЛОТИП: SDSM 2980 (AMNH cast 80132), левый верхнечелюстной фрагмент с M1 и альвеолами P4 и M2.
РАСПРОСТРАНЕНИЕ: уитний – ранний арикарий Северной Америки.
ОБСУЖДЕНИЕ: представляет собой одну из первых тенденций к гиперкарниворности в подсемействе Borophaginae. Отсутствие связи между метаконом и лингвальным цингулумом на M2 и отсутствием протостилида на m1 указывает только на базисную связь с флаоциноидной кладой, несмотря на то, что гипокон на M1 присутствует у всех форм.
Уонг (Wang, 1999) признаёт Oxetocyon и Otarocyon как два разных рода, хотя не исключает возможность, что они могут быть сестринскими таксонами.
OTAROCYON Green, 1954
РАСПРОСТРАНЕНИЕ: арикарий-орелланий Северной Америки
ДИАГНОЗ: виды Otarocyon объединяются многими синапоморфиями: гипертрофированная слуховая булла, сплющённый в передне-заднем направлении парокциптальный выступ, крупная надвисочная ямка, укороченный рострум, расширенная мозговая капсула, увеличенный протокон на P4, высокий лингвальный цингулум на M1, конический энтоконид m1, отделённая от метаконида глубокая выемка и пр.
ОБСУЖДЕНИЕ: явно специфический внешний вид Otarocyon без какого-либо очевидного предка представляет проблему филогенетической интерпретации. Несмотря на то, что Otarocyon разделяет с Archaeocyon прогрессивные характеристики борофагин (уменьшенный парастиль на M1, более массивный талонид m1 и высокий энтоконид на m2), эволюция, благодаря которой эти признаки были достигнуты, как кажется, предполагают независимое развитие этих символов.
При всей своей уникальной морфологии у Otarocyon есть живой аналог - фенек (Vulpes (Fennecus) zerda), разделяющий с Otarocyon множество прогрессивных признаков. В то же время фенек отличается от Otarocyon дополнительными деталями, сближающими последнего скорее с борофагинами.
Otarocyon macdonaldi
ГОЛОТИП: AMNH 38986 - череп с зубным рядом и нижней челюстью.
ОБСУЖДЕНИЕ: несмотря на фиксацию в ореллании и временной хиатус с арикийским видом O. cooki (по крайней мере, в 2 млн лет), O. macdonaldi в основном, демонстрирует все морфологические специализации, присутствующие у O. cooki. Другими словами - после того, как в процессе эволюции были получены основные адаптивные признаки, линия Otarocyon оставалась практически неизменной.
Otarocyon cooki 
Otarocyon cooki (FAM 49020)
ГОЛОТИП: SDSM 54308, фрагментарная правая нижнечелюстная ветвь с p4-m2.
ОБСУЖДЕНИЕ: более ранняя, арикарийская форма.
RHIZOCYON Wang,1999
Rhizocyon oregonensis (AMNH 6879), лектотип
ТИПОВОЙ ВИД: Rhizocyon oregonensis Merriam, 1906.
РАСПРОСТРАНЕНИЕ: ранний арикарий Северной Америки.
ЛЕКТОТИП: AMNH 6879 - череп со сравнительно полным зубным рядом.
ДИАГНОЗ: отличается от вышеописанных форм более широкой лобной областью, квадратными верхними молярами с переднерасширенным лингвальным цингулумом и связью лингвального цингулума с метаконулем на M2. Отличается от представителей трибы Phaoocyonini морфологией слуховой буллы, задненаправленным парокципитальным выступом, редуцированным метаконулем на M1 и отсутствием протостилида на нижних молярах.
ОБСУЖДЕНИЕ: ранее упоминался как Hesperocyon gregarious, позднее относился к Archaeocyon pavidus. Кладистический анализ предполагает, что Rhizocyon представляет собой базального борофагина (Wang, 1999).
Триба Phlaocyonini
РАСПРОСТРАНЕНИЕ: ранний арикарий – ранний барстовий (30-16 млн лет назад) Северной Америки (поздний олигоцен – средний миоцен).
ДИАГНОЗ: прогрессивные символы, выделяющие флаоционов, включают крупный метаконуль на M1 и наличие протостилида на m1. Также отличаются от Cormocyon наличием примитивного укороченного тригонида на m1.
CYNARCTOIDES McGrew, 1938
ТИПОВОЙ ВИД: Cynarctus acridens
ДИАГНОЗ: прогрессивные формы демонстрируют развитый рострум, длинные челюсти, узкие и длинные премоляры, конические и высокие бугорки на нижних коренных зубах, удлинённые вторые премоляры и пр.
ОБСУЖДЕНИЕ: все дискуссии о систематике Cynarctoides были, в основном, основаны на нескольких экземплярах и морфологические пробелы оставляли место для спекулятивных гипотез. В настоящее время имеется большое количество образцов в широком спектре географических и геологических локаций. Кроме того, описаны новые виды, которые в совокупности представляют собой почти непрерывную серию промежуточных ступеней развития к своеобразной форме, совершенно отличающейся как от Phlaocyon, так и от Cynarctus.
Cynarctoides lemur
ГОЛОТИП: AMNH 6888 - череп с альвеолами I1-C1 и рядом P1-M2.
ОБСУЖДЕНИЕ: протостилид на m1 традиционно использовался для дифференцирования Cynarctoides от современного ему Phlaocyon, однако более поздние наблюдения предполагают, что протостилид, по-видимому, присутствует у обоих видов. Таким образом, протостилид является синапоморфией клады.
Фрагментарные материалы, относящиеся к Phaoocyon achoros, по-видимому, относятся к этому же виду.
Cynarctoides roii 
Cynarctoides roii (SDSM 53321), голотип
ГОЛОТИП: SDSM 53321 (AMNH 129863) - частичная правая нижнечелюстная ветвь из Южной Дакоты.
ОБСУЖДЕНИЕ: очень мелкая борофагина. Её малый размер и примитивная морфология в сопоставлении с C. lemur и C. harlowi позволет предположить, что C. roii, C. harlowi и, возможно, C. lemur, образуют собственную небольшую кладу, в пределах которой уменьшение размера, по-видимому, является основной тенденцией.
Cynarctoides harlowi
ГОЛОТИП: ACM 31-34 (AMNH 48830) - частичная правая нижнечелюстная ветвь из Вайоминга.
ОБСУЖДЕНИЕ: борофагина небольшого размера. Высокая степень износа моляров и плохая сохранность голотипа делают определение его таксономического статуса сложным. Несмотря на то, что ACM 31-34 хорошо согласуется с некоторыми образцами C. roii, его гораздо более молодой возраст (поздний арикарий) и отличия в пропорциях нижних зубов предполагают валидность отдельного вида.
Cynarctoides luskensis
ГОЛОТИП: FAM 49005, частичный череп и фрагментарная мандибула из Вайоминга.
ОБСУЖДЕНИЕ: в настоящее время идентифицируется как переходная форма, демонстрирующая филогенетическую связь между прогрессивными видами Cynarctoides и примитивным C. lemur. Утоньшённая нижнечелюстная ветвь и структура нижних зубов отличаются от продвинутых Cynarctoides, тогда как верхние моляры сходны с C. lemur
Cynarctoides gawnae
ГОЛОТИП: FAM 49249, частичный череп с фрагментарной нижней челюстью из раннего хемингфордия Нью-Мексико.
ОБСУЖДЕНИЕ: в неопубликованной диссертации по геологии и палеонтологии Гауна (Gawne, 1973) описан новый вид Cynarctoides, который ранее никогда официально не публиковался. Данный таксон является примером переходной формы, соединяющей C. acridens, который обладает многими передовыми характеристиками рода, с C. luskensis. Эта комбинация переходных признаков позволяет понять эволюционные преобразования, проявляемые видами Cynarctoides в приобретении их своеобразной морфологии. Тот факт, что самые ранние находки C. acridens фиксируются в позднем арикарии Новой Мексики и Небраски, подтверждают, что C. gawnae, должно быть, также имел исторические корни в арикарии.
Cynarctoides acridens
ГОЛОТИП: AMNH 82558 (HC 144), фрагментарная правая нижнечелюстная ветвь из Небраски.
ОБСУЖДЕНИЕ: несмотря на почти одинаковый размер и форму голотипов C. acridens и C. mustelinus, Мэтью (Matthew, 1932) считал C. mustelinus отличающимся от первого и морфологически ближе к C. lemur. Однако недостаток материала в то время позволил Мэтью ориентироваться скорее на стратиграфию, потому что в настоящий момент ясно, что C. acridens имел почти непрерывный диапазон распространения от среднего арикария до раннего барстовия Великих равнин и юго-западных Соединенных Штатов как с точки зрения географического ареала, так и геологического распределения.
Cynarctoides emryi
ГОЛОТИП: UNSM 25455 (FAM 96711) - частичная правая нижнечелюстная ветвь из раннего хемингфордия Небраски.
ОБСУЖДЕНИЕ: сосуществовал с C. acridens, но интересно отметить, что более примитивные C. acridens пережили C. emryi и выжили в раннем барстовии, а также имели гораздо более широкий ареал зоогеографического распространения.
Как терминальный член рода, C. emryi является наиболее гипокарниворным из всех видов. Фактически, морфология зубов этого небольшого псового во многом вышла за пределы обычного диапазона гипокарниворного хищника и демонстрирует приспособления к насекомоядности или всеядности вследствие его довольно селенодонтных нижних коренных зубов. Подобная специализация чрезвычайно редко встречается среди представителей Carnivora и известна только у лофоциониновых вивверовых из европейского моцена.
PHLAOCYON Matthew, 1899
РАСПРОСТРАНЕНИЕ: ранний арикарий – поздний хемингфордий (поздний олигоцен – ранний миоцен) Северной Америки (30-16 млн лет назад).
ДИАГНОЗ: синапоморфии, которые объединяют представителей Phaoocyon, заключаются в наличии массивных премоляров, расширенных p4, квадратных и удлинённых верхних коренных зубов и широким талонидом на m1.
Phlaocyon minor 
Phlaocyon minor (F:AM 49054)
ГОЛОТИП: AMNH 12877 - частичные нижнечелюстные ветви из позднего арикария Южной Дакоты.
ОБСУЖДЕНИЕ: ранее образцы объединялись с Cynodesmus (Cook and Macdonald, 1962) и путаница продолжалась довольно длительное время. Теперь Cynodesmus признан принадлежащим Hesperocyoninae (Wang, 1994).
Phlaocyon latidens
ГОЛОТИП: AMNH 6896 - частичный череп с левой нижнечелюстной ветвью из раннего арикария Орегона.
ОБСУЖДЕНИЕ: на протяжении длительного времени со времени их первоначального описания Коупом (Cope, 1879), Cynarctoides lemur и Phlaocyon latidens рассматривались как близкие друг к другу виды. Действительно, их сходные размеры и многочисленные общие примитивные символы затрудняют дифференциацию множества фрагментарных образцов. В свете современной филогении признаётся ряд производных признаков у P. latidens, которые индивидуально могут выглядеть гомоплазийными или примитивными, но в значительной степени указывают на принадлежность к данному роду.
Phlaocyon annectens
ГОЛОТИП: CMNH 1602 (AMNH 89668), левые и правые верхнечелюстные фрагменты из позднего арикария Небраски.
ОБСУЖДЕНИЕ: Уонгом синонимизирован с P. willistoni (Wang, 1999). Общий размер и форма голотипа P. willistoni довольно близка с образцами P. annectens, однако этот образец имеет более крупный P4 с более выраженным протоконом и небольшим гипоконусом. По Уонгу, эти особенности находятся в пределах обычного диапазона видовых вариаций и представляют собой промежуточные стадии в тенденции к более продвинутым таксонам (например, P. leucosteus и P. marlandlandensis). Ещё один экземпляр относится к P. willistoni согласно размеру и массивности p3. Также имеются и другие образцы, интерпретируемые Петерсоном (Peterson, 1924) как принадлежащие к данному таксону, но в настоящее время материал вида P. willistoni разделён между Cynarctoides roii и P. annectens.
Phlaocyon achoros
Phlaocyon achoros (UF 18389), голотип
ОБСУЖДЕНИЕ: последний анализ показывает, по крайней мере, три таксона среди мелкоразмерных канид: Cynarctoides lemur, Phlaocyon achoros и Cormocyon copei (представленный M2), которые, как минимум, коэволюционно связаны с P. achoros. К тому же имеется образец, демонстрирующий повышенный парастиль на P4 и зарождающееся развитие дополнительного выступа между метаконулем и метаконом на M1 - здесь может возникнуть соблазн проследить эволюцию от achoros до multicuspus.
Phlaocyon multicuspus
ГОЛОТИП: FMNH UC1482 (AMNH 108069), частичный череп с альвеолами передних зубов и рядом щечных из позднего арикария Вайоминга.
ОБСУЖДЕНИЕ: МакГрей (McGrew, 1941) отметил, что «сходство в общей структуре черепа и основной схеме зубов между Phlaocyon и Aletocyon не оставляет сомнений в том, что эти два таксона тесно связаны». Исходя из предполагаемых морфологических расстояний, очевидно, что род Aletocyon может быть использован для представления небольшой клады, состоящей из родов achoros-multicuspus.
Phlaocyon leucosteus 
Phlaocyon leucosteus (AMNH 8768), голотип
ГОЛОТИП: AMNH 8768 - череп с нижней челюстью и фрагментарная посткраниаль из позднего арикария Колорадо.
ОБСУЖДЕНИЕ: см. общий диагноз для рода.
Реконструкция Phlaocyon leucosteus
Phlaocyon marslandensis
ГОЛОТИП: FMNH P26314 (AM5H 95585), левая частичная верхнечелюстная кость с почти полным зубным рядом из раннего хемингфордия Небраски
ОБСУЖДЕНИЕ: МакГрей (McGrew, 1941) предположил, что P. marslandensis является более прогрессивной формой P. leucosteus частично из-за предполагаемой разницы в возрасте между двумя видами. Однако такой связи в настоящее время противоречат аутопоморфии у P. leucosteus; расширенный p4 и более выраженный протостилид.
Phlaocyon yatkolai
ГОЛОТИП: UNSM 62546, правая нижнечелюстная ветвь с полным зубным рядом (и альвеолой m3) из раннего хемингфордия Небраски.
ОБСУЖДЕНИЕ: несмотря на отсутствие переходных форм между голотипами P. yatkolai и P. mariae (главным образом верхние зубы), налицо сестринская взаимосвязь этих двух видов.
Phlaocyon mariae
ГОЛОТИП: F: AM 25466, верхние и нижние зубы и краниальные фрагменты из раннего хемингфордия Небраски
ОБСУЖДЕНИЕ: плохая сохранность голотипа препятствует более полной оценке филогенетических отношений, но ясно, что это необычный таксон со смесью мозаичных признаков. Наличие гиперкарниворных признаков в преимущественно гипокарниворной кладе вызывает недоумение и вызывает сомнения относительно его филогенетического положения. Возможно, P. mariae демонстрирует пример крупноразмерной формы, вследствие этого ставшей более хищной.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#952 30 December 2018 13:18:43
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Триба Borophagini
ТИПОВОЙ ВИД: Borophagus Cope, 1892.
РАСПРОСТРАНЕНИЕ: арикарий – бланк Северной Америки
ДИАГНОЗ: в отличие от Phaoocyonini и других базальных форм, Borophagini демонстрируют прогрессивную характеристику удлинённого тригонида на m1. Также имеется ряд краниальных и дентальных синапоморфий, свидетельствующих о тенденции к мезо-гиперкарниворности, а также укрупнении размеров.
CORMOCYON Wang and Tedford, 1992
РАСПРОСТРАНЕНИЕ: арикарий Северной Америки (поздний олигоцен – ранний миоцен)
ОБСУЖДЕНИЕ: в одной статье Уонг и Тедфорд (Wang and Tedford, 1992) попытались прояснить концепцию Nothocyon , которая стала сборным таксоном для многих небольших ископаемых форм. Значительная часть прежней путаницы вокруг Nothocyon, по мнению Уонга (Wang, 1999) основана на неправильной интерпретации некоторых образцов, часть которых принадлежат париктинам (примитивным урсоидам), а других – борофагиновым канидам. Однако морфологическая разница между валидным Cormocyon и «мозаичным» Nothocyon в качестве базальных борофагин до сих пор не установлена. В настоящее время Cormocyon считается базальным парафилетическим родом трибы Borophagini.
Cormocyon haydeni 
Cormocyon haydeni (FAM 49448), голотип
ГОЛОТИП: FAM 49448 - череп с нижней челюстью и посткраниальным материалом из позднего арикария Южной Дакоты.
ОБСУЖДЕНИЕ: по-видимому, является наиболее базальным видом борофагин – линия мелкоразмерных псовых, отличающихся морфологией окципитального выступа и молярами с более увеличенными выступами.
Cormocyon copei
РАСПРОСТРАНЕНИЕ: ранний арикарий Орегона.
ОБСУЖДЕНИЕ: нынешняя концепция C. copei, в основном, основана на ранних образцах ввиду морфологии зубов, позволяющих отнести их к примитивным борофагинам с такими характеристиками, как укороченный Р4, выраженные метаконули М1-М2 и др. В то же время некоторые дентальные признаки могут указывать на отдельный вид (или даже сестринский род), который выглядит прогрессивнее, чем C. haydeni.
DESMOCYON Wang,1999
РАСПРОСТРАНЕНИЕ: - поздний арикарий – ранний хемингфордий Северной Америки
ДИАГНОЗ: в сравнении с Cynarctina у Desmocyon отсутствуют гипокарниворные признаки: видная возвышенный мандибулярный мыщелок, укороченный P4, протостилид на m1, широкий талонид на m1 и крупные m2. Desmocyon примитивен по сравнению с Metatomarctus тем, что I3 не увеличен и не имеет хорошо развитого латерального бугорка, а P4 имеет только слабый передний гребень на параконе.
Desmocyon thomsoni 
Desmocyon thomsoni (AMNH 12874), голотип
ГОЛОТИП: AMNH 12874 - череп с нижней челюстью и посткраниальные фрагменты, поздний арикарий.
ОБСУЖДЕНИЕ: первоначально приписывали к Cynodesmus, а позже интерпретировали как связующий таксон между Hesperocyon и Tomarctus. Морфо-филогенетический анализ Уонга (Wang, 1999) показывает, что данный таксон не имеет отношения к канинам, а скорее связан с серией борофагинных псовых.
Некоторые особи D. thomsoni из Небраски (например, UNSM 26550 и 26664) заметно меньше, чем из Вайоминга. Подобное различие в размерах в целом выходит за пределы полового диморфизма но, не имея никаких морфологических признаков для разделения этих образцов, следует учитывать возможность уменьшения размера в пределах вида.
Desmocyon matthewi
ГОЛОТИП: FAM 49177 - полный череп и нижняя челюсть с фрагментарной посткраниалью.
ОБСУЖДЕНИЕ: образцы D. matthewi довольно редки. Очевидно, филогенетически быстро привели к появлению Cynarctina и Metatomarctus.
Подтриба CYNARCTINA
РАСПОСТРАНЕНИЕ: ранний хемингфордий – ранний кланендорий Северной Америки.
ДИАГНОЗ: Cynarctina разделяет с Metatomarctus единственную прогрессивную характеристику (как кажется) - боковая дополнительная вершинка на I3, отличающая его от более примитивного Desmocyon. Подтриба Cynarctina отличается от Metatomarctus и Euoplocyon гипокарниворными характеристиками: морфологией нижней челюсти, укорочением P4 относительно верхних моляров, увеличенный M2, протостилид на m1, удлинённые m2 и прочее.
PARACYNARCTUS Wang,1999
РАСПРОСТРАНЕНИЕ: ранний хемингфордий – поздний барстовий
ОБСУЖДЕНИЕ: отсутствие выраженных признаков гиперкарниворности у Paracynarctus, проявлявшиеся уже на умеренном уровне, отделяет его от Cynarctus. Более того, Paracynarctus отнюдь не обладает многочисленными синапоморфиями, присущими цинарктинам, которые разделяют с ним редукцию премоляров, усложнение морфологии нижних коренных зубов, отличияем в слуховой области и другие особенности. По большому счёту, есть немало как общих, так и разнящихся характеристик между Paracynarctus и Cynarctus, которые предполагают скорее сестринское отношение между ними.
Paracynarctus kelloggi
ГОЛОТИП: UCMP 11562 (AMNH 27873), правая и левая фрагментарные нижнечелюстные ветви из Невады.
ОБСУЖДЕНИЕ: предыдущие исследования размещали этот таксон в Tomarctus и Tephrocyon. По мнению Уонга, морфологически и хронологически P. kelgloggi, по-видимому, не согласуется с другими видами Tomarctus а сочетает, в более широком представлении, их мезо- и гипокарниворные признаки (хотя бы эмальная сниженность премоляров). Некоторые особенности хищнических зубов предполагают родство Paracynarctus с цинарктинам, хотя ограниченный фоссильный материал пока что ставит под сомнение надёжную идентификацию.
Paracynarctus sinclairi 
Paracynarctus sinclairi (FAM 61009), голотип
ГОЛОТИП: FAM 61009 с практически полным зубным рядом из раннего барстовия Небраски.
ОБСУЖДЕНИЕ: умеренно гиперкарниворная характеристика зубного ряда демонстрирует совершенно иную конфигурацию по сравнению с Cynarctus (форма гипокона P4, увеличенный лингвальный цингулум на M1 наряду со сниженным гипоконом, что не наблюдается у Cynarctus). Явно, что это форма, чётко отличающаяся от Cynarctus (но интересно бы определить её отношения с Metatomarctus и Desmocyon).
CYNARCTUS Matthew, 1902
РАСПРОСТРАНЕНИЕ: ранний барстовий - поздний кларендонский Небраски, Южной Дакоты и Техаса.
ДИАГНОЗ: прогрессивные символы, отличающие Cynarctus от Paracynarctus, представляют собой некотоые отличия в слуховой булле, поперечно узкий и субквадратный M1, сильно изогнутый c1 и пр.
ОБСУЖДЕНИЕ: из нескольких независимых попыток борофагин в гипокарниворности Cynarctus является наиболее прогрессивным. По сути, данный таксон является довольно урсидным в плане уменьшения премоляров, наружного расширения M2, увеличении талонида на m1. Мэтью (Matthew, 1902) изначально считал, что Cynarctus – амфицидонид. Но морфология Cynarctus демонстрирует топологию среднего уха (увеличенный энтотимпаник, наличие алисфеноидного канала и др.) в пределах борофагинной клады.
Cynarctus galushai
ГОЛОТИП: FAM 27543, фрагментарный череп с частичной ветвью нижней челюсти из раннего барстовия Калифорнии.
ОБСУЖДЕНИЕ: представляет собой раннюю стадию специализированных гипокарниворных форм в линии Cynarctus. Признаки включают наличие узкого внешнего прохода, сужения M1, морфологию p4 и m1- эти признаки с одной стороны позволяют дифференцировать мезокарнивовидную линию, близкой к Metatomarctus, но всё же остаётся довольно большой морфологический разрыв между C. galushai и прогрессивной морфологией в C. saxatilis (через C. crucidens).
? Cynarctus marylandica
ГОЛОТИП: USNM 15561 (AMNH 48836), левая ветвь нижней челюсти с эрозированными молярами из раннего барстовия Мэриленда.
ОБСУЖДЕНИЕ: Тедфорд и Хантер (Tedford and Hunter, 1984) предположили, что материал из Мэриленда может представлять примитивного Cynarctus. Морфологически данные образцы не позволяют уверенно определить таксономическое положение этой формы, хотя их метрика позволяет разместить их в диапазоне среднеразмерных борофагин. Также об отличительности данного таксона может свидетельствовать некоторые морфологические отличия в m1.
Cynarctus saxatilis
ГОЛОТИП: AMNH 9453, правая и левая нижнечелюстные ветви с альвеолами i1-i3, р2 и c1-m3 из позднего барстовия Колорадо.
ОБСУЖДЕНИЕ: морфология нижних зубов голотипа C. saxatilis очень близка с C. crucidens. Однако в приписываемом материале отсутствуют экстремальные гипокариноподобные признаки, имеющиеся у C. crucidens – усиление цингулума верхнего моляра, удлинение M2 и постериорная позиция гипокона на этом зубе. Не исключено, что образцы включают форму видового ранга более примитивную, чем C. saxatilis.
Между saxatilis и galushai существует большая морфологическая разница, что отражено многочисленными апоморфными символами, всё же разделяющими два вида.
Cynarctus voorhiesi
ГОЛОТИП: FAM 105094, правая частичная нижнечелюстная ветвь из позднего барстовия Небраски.
ОБСУЖДЕНИЕ: несмотря на то, что C. voorhiesi слабо представлен ископаемым материалом, он заполняет морфологический разрыв между C. saxatilis и C. crucidens. Это почти точно промежуточное звено, как по размеру и форме, так и стратиграфически между этими видами.
Cynarctus crucidens
ГОЛОТИП: UNSM 25465 (AMNH 14307), правая частичная нижнечелюстная ветвь из раннего кларендония Небраски.
ОБСУЖДЕНИЕ: наличествует тенденция к уменьшению размера от более ранних видов Cynarctus до финальных. Параллельно с этим наблюдается редукция премоляров (особенно верхних).
Далквист (Dalquest, 1962) описывал фрагментарный череп (FMNH P25537), который был отнесён к C. fortidens, однако относимый разными исследователями к crucidens или к saxatilis. Тем не менее, некоторые диагностические признаки выглядят отличительными между C. fortidens и C. saxatilis.
METATOMARCTUS Wang,1999
РАСПРОСТРАНЕНИЕ: хемингфордий Северной Америки.
ДИАГНОЗ: прогрессивные символы, отличающие Metatomarctus от Desmocyon - I3 с одним боковым выступом (как у Cynarctina) и начальным развитием переднего гребня на P4. Metatomarctus примитивен по отношению к Euoplocyon, Psalidocyon и более высоким таксонам и не имеет таких продвинутых признаков, как высокая позиция нижнечелюстного мыщелка, редукция премоляров, укороченный P4, небольшой протостилид на m1, широкий талонид на m1 и удлинённые m2.
Metatomarctus canavus 
Metatomarctus canavus (UFV- 5260), голотип
ГОЛОТИП: правая частичная нижнечелюстная ветвь из хемингфордия Флориды.
ОБСУЖДЕНИЕ: изначально наличествовало шесть номинальных видов для канид из типового местонахождения, сузившееся до двух таксонов. Уонг (Wang, 1994) предлагает номинативное именование Tomarctus, исходя из фундаментальных различий между геспероционами и борофагиновыми псовыми. Позже гомологичный материал был обнаружен, помимо Флориды, и в Небраске, что является свидетельством широкого зоогеографического распределения этой формы.
EUOPLOCYON Matthew, 1924
Euoplocyon brachygnathus (CMNH 752), голотип
РАСПРОСТРАНЕНИЕ: ранний хемингфордий - ранний барстовий Северной Америки
ДИАГНОЗ: единственная прогрессивная характеристика, которая отличает Euoplocyon от Metatomarctus и более примитивных форм - наличие двух боковых выступов на i3. Euoplocyon не имеет увеличенного парастиля на P4, который присутствует у Psalidocyon и других псовых.
ОБСУЖДЕНИЕ: среди среднеразмерных борафагин Euoplocyon является наиболее гиперкарниворным в его более компактном нижнем карнассиале. Тедфорд и Фрэйли (Tedford and Frailey, 1976) первыми указали на возможную связь Euoplocyon с такими продвинутыми борофагами, как Tomarctus и Aelurodon, основываясь главным образом на дентальном сходстве. Такие структурные символы предполагают относительно базисную позицию в филогенезе - примерно вблизи Psalidocyon и Microtomarctus. В сущности, гиперкарниворная морфология Euoplocyon появляется в палеолетописи довольно внезапно без промежуточных форм.
Euoplocyon spissidens
ГОЛОТИП: MCZ 4246 (AMNH 56015), левая частичная нижнечелюстная ветвь из хемингфордия Флориды.
ОБСУЖДЕНИЕ: если spissidens правильно диагностирован как примитивный вид этого рода, то следует, что расширенное нёбо и связанные с ним особенности E. brachygnathus, должно быть, сформировались внутри рода Euoplocyon, независимо от Aelurodon и Epycyon.
Euoplocyon brachygnathus
ГОЛОТИП: CMNH 752 (AMNH 101208), левая частичная нижнечелюстная ветвь из раннего барствия Монтаны.
ОБСУЖДЕНИЕ: в качестве терминального представителя небольшой, но довольно гиперкарноподобной клады, E.brachygnathus отображает несколько черт, сходных с Aelurodon. В частности, в уменьшении вторых коренных зубов, а также общей тенденцией к редукции метаконуля на M1 и сужению талонида m1.
PSALIDOCYON Wang,1999
Psalidocyon marianae (FAM 27397), голотип
РАСПРОСТРАНЕНИЕ: ранний барстовий из Нью-Мексико и Небраски.
ДИАГНОЗ: символы, уникальные для этого рода, являются высокоразвитыми и усложнёнными: крупный клык с сжатой и вогнутой лингвальной поверхностью, усложнённые P1-P3 и p1-p4, открытый тригонид на m1 и пр.
Psalidocyon marianae
ГОЛОТИП: FAM 27397, череп с нижней челюстью из раннего барствия Нью-Мексико.
ОБСУЖДЕНИЕ: череп и верхние моляры Psalidocyon довольно типичны для мезокарниворных борофагов среднего размера, мало чем отличаясь от таковых у Desmocyon и Metatomarctus. Эта комбинация примитивных и производных признаков рамещает его выше Metatomarctus и ниже Microtomarctus.
MICROTOMARCTUS Wang,1999
Microtomarctus conferta (AMNH 17203), голотип
ДИАГНОЗ: признаки, уникальные для этого рода, представляют собой небольшие размеры, брахицефальный череп и укороченные клыки.
Microtomarctus conferta
ГОЛОТИП: AMNH 17203, правая ветвь из раннего барстовия Небраски.
ОБСУЖДЕНИЕ: небольшой размер M. conferta интерпретируется как результат увеличения размера в других морфологически сходных группах.
PROTOMARCTUS Wang,1999
Protomarctus optatus (AMNH 18916), голотип
ДИАГНОЗ: Protomarctus прогрессивен в отношении Microtomarctus и более примитивных таксонов в морфологии черепной коробки, но примитивней Terocrocyon, Aelurodontina и Borophagina дентальными особенностями.
Protomarctus optatus
ГОЛОТИП: AMNH 18916, нижняя челюсть из позднего хемингфордия Небраски.
ОБСУЖДЕНИЕ: обладает комбинацией признаков, чтобы быть предковым для клад Aelurodontina и Borophagina. Мэтью (Matthew, 1924) предположил, что Р. optatus может быть примитивной морфологической аберрацией Tomarctus hippophaga, но предпочёл сохранить именования в зависимости от дальнейших находок.
TEPHROCYON Wang,1999
Tephrocyon rurestris (UO 23077), голотип
РАСПРОСТРАНЕНИЕ: ранний барстовий Орегона и Техаса.
ДИАГНОЗ: прогрессивней по сравнению с Protomarctus более крупными размерами, более широким нёбом и удлинённым m2. Отличается от Aelurodontina небольшими премоларами, сниженным параконом на M1 и другими дентальныи особенностями.
ОБСУЖДЕНИЕ: вскоре после описания Мерриамом (Merriam, 1906) в него включили несколько средних таксонов среднего размера и Tephrocyon быстро стал таксономической мусорной корзиной для многих форм, однако после ревизии рода Canis (Matthew, 1930) был оставлен только типовой вид T. rurestris.
Tephrocyon rurestris
ГОЛОТИП: UO 23077 (AMNH 97225), череп с нижней челюстью из раннего барстовия Орегона.
ОБСУЖДЕНИЕ: В качестве геноголотипа Т. rurestris сыграл важную роль в формировании ранней концепции филогенеза канид. Хотя Уонг (Wang, 1994) помещает T. rurestris в трихотомию с трибами Aelurodontina и Borophagina, однако rurestris не имеет расширенных премоляров, которые являются производной характеристикой.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#953 30 December 2018 13:28:48
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Подтриба Aelurodontina
РАСПРОСТРАНЕНИЕ: ранний барстовий – поздний кларендоний (средний миоцен) Северной Америки (16-10 млн л.н.).
ДИАГНОЗ: всех представителей Aelurodontina объединяет синапоморфная особенность: равномерно увеличенные нижние премоляры с развитыми дополнительными бугорками. Продвинутые члены этой клады дополнительно демонстрируют производные такие характеристики, как расширенное нёбо, высокий сагиттальный гребень, сжатый латерально затылочный гребень и целый реестр дентальных символов. В то же время у них обычно отсутствуют производные признаки трибы Borophagina.
TOMARCTUS Cope, 1873
РАСПРОСТРАНЕНИЕ: барстовий Северной Америки.
ДИАГНОЗ: единственный прогрессивный признак, который объединяет Tomarctus и Aelurodon - расширенные премоляры. Также отличается от Protomarctus большими размерами, более широкой лобной частью; от Aelurodon сниженным сагиттальным гребнем уменьшенной слуховой буллой и рядом дентальных характеристик.
ОБСУЖДЕНИЕ: Tomarctus является одним из самых запутанных таксонов в исследованиях третичных канид. Эта путаница возникла из ранних филогенетических синтезов с Canidae (Matthew, 1930), которые следовали схеме с двумя дихотомическими линиями канид и схемы Мэтью придерживалось большинство последующих исследователей. Из филогенетического анализа Уонга (Wang, 1994) ясно, что даже допуская горизонтальную (градационную) классификацию, некоторые виды, относящиеся к Tomarctus (sensu lato), занимают базальные позиции в различных кладах борофагин и далеки от любых Caninае. Кладистически Tomarctus выглядит парафилетическим родом и относится только к трибе Aelurodontina.
Tomarctus hippophaga 
Tomarctus hippophaga (FAM 61156)
ГОЛОТИП: AMNH 13836, правая нижнечелюстная ветвь из раннего барстовия Небраски.
ОБСУЖДЕНИЕ: после ревизии Мэтью (Matthew, 1924) Т. hippophaga почти исчез из литературы, хотя Мэтью всё ещё питал некоторые сомнения в отношении этого решения. Дело в том, что наличествует некоторая разница в размерах голотипов T. brevirostris и T. hippophaga, а также разница в материале из некоторых местонахождений с образцами T. brevirostris, которые предполагают, что там наличествует более одного вида. Таким образом, восстановление этого таксона Уонгом является попыткой объяснить данные количественные и качественные различия.
Tomarctus brevirostris
ГОЛОТИП: AMNH 8302, правый нижнечелюстной фрагмент из раннего барстовия Колорадо.
ОБСУЖДЕНИЕ: до тех пор, пока Мэтью (1924) не описал полный образец, валидность данного таксона игнорировалась некоторыми исследованиями. Этот материал сыграл важную роль в позиционировании T. brevirostris в качестве примитивного предка почти для всех поздних борофаговых.
AELURODON Leidy, 1858
Aelurodon asthenostylus (LACM-CIT 781), голотип
РАСПРОСТРАНЕНИЕ: поздний кларендоний - ранний барстовий Северной Америки.
ДИАГНОЗ: виды Aelurodon обладают многими производными признаками, которые отличаются от Tomarctus: высокий сагиттальный гребень, увеличенная булла, увеличенный мастоидный отросток, увеличенные парастиль на p4, уменьшение защёчных зубов и пр.
ОБСУЖДЕНИЕ: очень робустные череп и зубы Aelurodon, во многом сходные с черепно-зубными адаптациями костнодробящих гиеновых, всегда привлекали внимание палеонтологов. Это привело к высокому числу видовых именований, впоследствии подвергавшихся неоднократной ревизии.
Aelurodon asthenostylus
ГОЛОТИП: LACM-CIT 781 (LACM-CIT 2816), частичный краниум и фрагментарная нижняя челюсть из барстовия Невады.
ОБСУЖДЕНИЕ: большинство прогрессивных признаков, которыми обладает A. asthenostylus, относятся к традиционной концепции рода. По-видимому, он постепенно эволюционировал до A. ferox и, если это так, разделение этих двух форм отражает скорее анагенетическое, а не кладогенетическое событие. С другой стороны, налицо большая морфологическая разница между Tomarctus brevirostris и A. asthenostylus.
Aelurodon mcgrewi 
Aelurodon mcgrewi (FAM 22410), голотип
ГОЛОТИП: AMNH 22410, фрагментарный череп с левой нижнечелюстной ветвью из позднего барстовия Небраски.
ОБСУЖДЕНИЕ: по мнению Уонга, образцы из Railway quarries, ранее приписываемые A. stirtoni, представляют собой примитивную форму - A. mcgrewi, которая преодолевает морфологический разрыв между A. stirtoni и другими видами.
Aelurodon stirtoni 
ГОЛОТИП: UCMP 33473, левая фрагментарная нижнечелюстная ветвь из позднего барстовия Небраски.
ОБСУЖДЕНИЕ: своеобразная морфология М1 первоначально привела Уэбба (Webb, 1969) даже к обозначению нового рода Strobodon. M1 настолько уникально сформирован среди Borophagini, что даже в с сравнении с голотиповыми образцами новый род выглядел вполне валидным. 
Скелет Strobodon
Теперь имеется дифферентность клады A. mcgrewi-stirtoni, связанной с кладой A. ferox-taxoides. Среди ограниченного материала кластера A. mcgrewi-stirtoni наблюдается довольно отчётливая морфологическая трансформация от A. mcgrewi до A. stirtoni. В то же время геологически более ранний A. mcgrewi ближе по морфологии к A. asthenostylus и гипотетическому предку A. ferox.
Относительно короткий хронологический промежуток в датировке находок подразумевает, что морфологическая трансформация от A.mcgrewi до A. stirtoni, очевидно, произошла быстро - менее чем за 500 000 тлн.
Aelurodon ferox
[url=https://radikal.ru] [/url
[/url
ГОЛОТИП: USNM 523 (AMNH 9960), правый изолированный P4.
ОБСУЖДЕНИЕ: изолированный P4 USNM 523 - голотип A. ferox, был источником разочарования для многих палеонтологов, причём даже затрагивался вопрос о валидности Aelurodon (Baskin, 1980). Такая неопределённость в отношении голотипа, возможно, была стимулирована общим нежеланием большинства исследователей отнести обширный объём образцов к этому виду.
Несмотря на отсутствие чёткой концепции видовой типизации, Aelurodon столь широко использовалось, что даже авторы, которые рассматривали голотип как специфически детерминированные, не захотели отказаться от A. ferox, чтобы иметь возможность продолжить использование родового именования. В свете филогенетического исследования Уонга диагноз Баскина (Baskin, 1980) остается наиболее достоверной характеристикой крупных гиеноподобных борофагов. Тем не менее, многие спорные вопросы по-прежнему остаются в силе – выглядит так, что решение сохранить A. ferox в отличие от A. taxoides выглядит несколько произвольным и призвано лишь обозначить эволюционную стабильность.
Aelurodon taxoides 
ГОЛОТИП: YPM-PU 10635 (AMNH 97777), левая нижнечелюстная ветвь из кларендония Небраски.
ОБСУЖДЕНИЕ: в качестве конечного представителя линии к гиеноподобным адаптациям A. taxoides обладает очень выраженной кранио-дентальной морфологией. Его крупные размеры и массивные премоляры позволяют легко распознать эту форму. Таким образом, относительно хорошо сохранившийся голотип позволяет избежать таксономических сомнений, связанных с другими видами Aelurodon.
Отличительная морфология поляризует эту группу с массивными премолярами за счёт уменьшения моляров (например, редукция тригонида на m1). Противоположная характеристика, как правило, присуща группе Epicyon-Borophagus, у которых передние премоляры значительно уменьшены, но моляры остаются практически нередуцированными.
Подтриба Borophagina
ТИПОВОЙ ВИД: Borophagus Cope, 1892.
РАСПРОСТРАНЕНИЕ: поздний хемингфордий – поздний бланк (ранний миоцен-ранний плейстоцен) Северной Америки (16-1,8 млн л.н.).
ДИАГНОЗ: прогрессивные признаки, объединяющие Borophagina как монофилетическую группу, включают дорсально увеличенную куполообразную лобную область, редукцию или отсутствие цингулумных бугорков на премолярах и протокона на Р4. Обычно у Borophagina отсутствуют такие производные признаки Aelurodontina как высокий сагиттальный гребень, боковые бугорки на резцах, увеличение премоляров и пр.
PARATOMARCTUS Wang,1999
Paratomarctus temerarius (USNM 768), голотип
ТИПОВОЙ ВИД: Paratomarctus euthos McGrew, 1935.
РАСПРОСТРАНЕНИЕ: поздний хемингфордий - поздний кларендоний
ДИАГНОЗ: отличается от Aelurodontina следующими производными признаками: куполообразный лоб, уменьшенные или отсутствующие цингулумные бугорки и др. Отличается от кластера Carpocyon-Borophagus следующими примитивными характеристиками: булла небольшая, парастиль на P4 не увеличен. Аутапоморфия заключается в равномерном уменьшении премоляров.
Paratomarctus temerarius
ЛЕКТОТИП: USNM 768, правый нижнечелюстной фрагмент с m1 из позднего барстовия Небраски.
ОБСУЖДЕНИЕ: лектотип (два каталогизированных образца, из которых USNM 768 был избран лектотипом) и приписываемые к этому таксону образцы из других местонахождений все ещё оставляют место для дискуссий о дивергенции данного вида на две формы - каноничную Р. temerarius и Р. optatu как более раннюю.
Paratomarctus euthos
ГОЛОТИП: UCMP 29282, череп с альвеолами I1-I2 и I3-M2 из позднего барстовия Небраски.
ОБСУЖДЕНИЕ: Р. euthos изначально позиционировался как форма, ведущая непосредственно к Canis (McGrew, 1935). Впоследствии также немало споров вызвала диагностика образцов, представляющих собой несвязанные гнато-краниальные фрагменты из разных местонахождений. В настоящее время по мнению Уонга (Wang, 1994), отсутствие аутоапоморфий у P. temerarius и стратиграфическая непрерывность между P. temerarius и P. euthos указывают на линейную связь между этими таксонами.
CARPOCYON Webb, 1969
РАСПРОСТРАНЕНИЕ: ранний барстовий - поздний хемфилий (16-5 млн) Северной Америки.
ДИАГНОЗ: некоторые синапоморфии объединяют Carpocyon с кладой Epicyon-Borophagus и отличают его от Paratomarctus и более примитивных таксонов: увеличенная булла, укороченность наружного слухового прохода, боковые бугорки на I3, уменьшенные дополнительные бугорки и увеличенный парастиль на p4.
ОБСУЖДЕНИЕ: имеет некоторое сходство с Cynarctus из-за сходных адаптаций к гипокарниворности. В анализе Уонга он отличается от Cynarctus наличием заметного куполообразного лба, выраженного парастиля на P4 и более массивного p4 - характеристик, которые указывают на связь с кластером Protepicyon-Borophagus.
Carpocyon compressus
ГОЛОТИП: AMNH 8543, левая фрагментарная нижнечелюстная ветвь из позднего барстовия Небраски.
ОБСУЖДЕНИЕ: таксономическое обоснование Уонга обосновано филогенетическим анализом, который предусматривает две переходных формы, C. webbi и C. robustus, связывающие начальных и терминальных представителей рода.
Carpocyon webbi 
Carpocyon webbi (FAM 61328), голотип
ГОЛОТИП: FAM 61328, череп из позднего барстовия Небраски.
ОБСУЖДЕНИЕ: C. webbi достигает наибольшего размера из представителей рода Carpocyon и, в среднем, дентальная морфометрия у C. webbi на 16% больше, чем у других видов. В то же время аналогия между образцами из Нью-Мексико и Небраски остаётся сомнительной.
Carpocyon robustus
ГОЛОТИП: UCMP 33569 (AMNH 80158), фрагментарный скелет с частичными нижнечелюстными ветвями из кларендония Калифорнии.
ОБСУЖДЕНИЕ: филогенетический анализ предполагает, что C. robustus, возможно, эволюционировал независимо от основной линии Carpocyon. Не исключено, что столкнувшись с такими конкурентными крупноразмерными и более многочисленными хищниками как Epicyon и Aelurodon, C. robustus снизил размер в сравнении с предковой формой C. webbi, трофически «переключившись» на грызунов и зайцеобразных.
Carpocyon limosus
ГОЛОТИП: UF 12069 (AMNH 97775), два верхнечелюстных фрагмента из позднего хемфиллия Флориды.
ОБСУЖДЕНИЕ. ввиду ограниченного хроногеографического ареала описание данного таксона неполное и, возможно, является финальным представителем гипокарниворной ветви борофагин.
PROTEPICYON Wang,1999
РАСПРОСТРАНЕНИЕ: поздний барстовий Северной Америки.
ДИАГНОЗ: в отличие от Carpocyon и Paratomarctus, прогрессивные признаки, которые Protepicyon разделяет с кладой Epicyon-Borophaus, включают высококонечный паракон на M1 и широкий p4 относительно p2-p3. Примитивные признаки, отличающие Protepicyon от Epicyon: меньшая редукция премоляров, дифферентность протокона и парастиля Р4 и пр.
Protepicyon raki 
Protepicyon raki (FAM 61705), голотип
ГОЛОТИП: FAM 61705, фрагментарный череп с частичной правой нижнечелюстной ветвью из барстовия Калифорнии.
ОБСУЖДЕНИЕ: таксон имеет короткий хроногеографический диапазон, совпадая с Aelurodon asthenostylus. При этом оба вида имеют примерно одинаковый размер и трудноразличимы при фрагментарности образцов. Как промежуточная форма, Protepicyon является предком Epycon.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#954 30 December 2018 13:44:16
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
EPICYON Leidy, 1858
Epicyon saevus и Е. haydeni
РАСПРОСТРАНЕНИЕ: ранний кларендоний - поздний хемфиллий (средний миоцен - плиоцен) Северной Америки (13-5 млн).
ДИАГНОЗ: синапоморфии, объединяющие Epicyon с кластером Borophagus - более широкое нёбо, латеральные бугорки на I3 плохо выражены, премоляры редуцированы, протокон и парастиль на Р4 соединены гребнем, внешний цингулум на параконе M1 отсутствует и пр. Эпицион отличается от борофага следующими примитивными чертами: более удлинённая лицевая область; не столь увеличенная лобная часть черепа; P1-P3 и p1-p3 относительно меньше P4 и p4; задние бугорки на P2-P3 и p2-p3 не уменьшены; протокон на P4 снижен; p4 более узкий по сравнению с другими премолярами; большеберцовая кость более тонкая и удлинённая по отношению к голени 90% (против 100% у борофага) и др.
ОБСУЖДЕНИЕ: Epycon первоначально описан Лейди (Leidy, 1858) как подрод Canis, но вскоре признан валидным, хотя типовой вид был помещен в Aelurodon. Позже эпицион был восстановлен как отдельный таксон Баскиным (Baskin, 1980), что подтверждается более поздними филогенетическими анализами.
В то же время между терминальными родами борофагин существует значительная разница в дентальной морфологии. Фокус наибольшего приложения усилий у Epycon находится в паре P4/p4 (как и у Borophagus), тогда как у Aelurodon, как представляется, более широкий диапазон - P3/p3 и P2/p2 (Werdelin, 1989).
Epicyon aelurodontoides 
Epicyon aelurodontoides (FAM 67025), голотип
ГОЛОТИП: F AM 67025, фрагментарный череп из раннего хемфиллия Канзаса.
ОБСУЖДЕНИЕ: этот вид обладает мозаичной смесью черепных и дентальных особенностей Aelurodon и Epicyon - такие характерные для Aelurodon признаки, как высокий сагиттальный гребень, постериорно спроецированный затылочный, редуцированный внутренний цингулум на M1 и очень уменьшенный M2. С другой стороны, FAM 67025 демонстрирует сходство с кладой Epicyon-Borophagus ввиду наличия прогрессивных признаков - слегка куполообразный лоб, уменьшенный внешний цингулум M1 и небольшие премоляры.
В частности, редукция премоляров является уникальным признаком, не наблюдаемым у Aelurodon, который имеет противоположную тенденцию к их укрупнению. Также наличествуют и другие особенности, свидетельствующие о морфологическом тождестве клады Epicyon-Borophagus.
Epicyon saevus 
Epicyon saevus. FAM 67331
ГОЛОТИП: USNM 126 (AMNH 97243), левая фрагментарная нижнечелюстная ветвь из раннего кларендония Небраски.
ОБСУЖДЕНИЕ: эволюционное появление E. saevus до сих пор дискуссионно. Недавнее исследование Манта (Munthe, 1989), рассматривавшего посткраниаль E. saevus, предполагает, что он (как и E. haydeni, который находится в том же размерном диапазоне), имел относительно тонкие конечности и, вероятно, предпочитал более открытые ландашфты. Т. е. он был курсориальным хищником и, судя по краниодентальной морфологии, являлся активным хищником - возможно, склонным к трупофагии.
Epicyon haydeni 
ГОЛОТИП: USNM 127 (AMNH 97244), правая нижнечелюстная правая ветвь из позднего кларендония Небраски.
ОБСУЖДЕНИЕ: обилие образцов, крупные размеры, широко распространенное географическое распределение и длительная хронология в совокупности обеспечивают благодатную почву для таксономических проблем в отношении E. haydeni.
Учитывая обширный стратиграфический контекст, общий материал значительно шире, чем отдельный вид – возможно, E. mortifer, синонимизированный с E. haydeni, является позднекларендонским подвидом. Объективно, Уонг (Wang, 1994) признаёт две морфологически гомологичных, но разноразмерных группы, которые наличествовали в течение позднего кларендония и раннего хемфиллия. С учётом присутствия в это же время крупного Aelurodon и небольшого Borophagus littoralis.
Хотя различия в размерах между E. saevus и E. haydeni можно объяснить половым диморфизмом, Уонг рассматривает их как разные виды по тем причинам, что никакие современные псовые не демонстрируют такие большие отличия в размерах между самцами и самками как в морфометрии, так и в кранио-дентальных пропорциях.
BOROPHAGUS Cope, 1892
ТИПОВОЙ ВИД: Borophagus diversidens Cope, 1892.
ДИАГНОЗ: синапоморфии, объединяющие формы Borophagus, проявляются в высококупольности лобной области; укороченной лицевой части, крупный p4 в сравнении с другими премолярами и хищническими зубами, почти равным или превышающими высоту тригонида m1 и пр.
ОБСУЖДЕНИЕ: исследование Манта (Munthe, 1979) показывает, что у борофага (в сравнении с Osteoborus) - самые короткие конечности среди борофагин, основываясь на посткраниальном материале B. secundus и B. diversidens как наиболее высокоразвитых форм. Независимо от того, подтверждают ли подобные локомоторные отличия меньшую курсориальность для Borophagus, для данных предположений необходим более скрупулёзный анализ посткраниальных скелетов Epicyon и Borophagus.
Borophagus littoralis
ГОЛОТИП: UCMP 31503 (AMNH 30073), почти полный череп из раннего кларендония Калифорнии.
ОБСУЖДЕНИЕ: Borophagus littoralis и, возможно, синонимизируемый с ним B. diabloensis, представляют собой самую примитивную форму в кластере [i]Borophagus (Vander Hoof, 1931; Richey, 1938) - данный вывод вполне поддерживается Уонгом.
Borophagus pugnator
ГОЛОТИП: DMNH 184 (AMNH 113864), передняя часть черепа с неполным зубным рядом из хемфиллия Колорадо.
ОБСУЖДЕНИЕ: данный таксон либо игнорировался, либо лишь поверхностно упоминался в исследованиях линии Osteoborus - Borophagus. Таким образом, такие виды, как B. galushai, были установлены главным образом путём демонстрации отличий от O. cyonoides – по сути, B. galushai находился на той же эволюционной стадии, что и B. pugnator[/i].
Ископаемый материал очень обилен. Меньшие образцы из Канзаса морфологически примерно одинаковы на эволюционном этапе с образцами из Колорадо, но согласно филогенезу Уонга, более ранний B. littoralis является сестринским таксоном для B. pugnator, а мелкие особи из Swayze Quarry представляют собой популяционное уменьшение размера. Является ли это прогрессивностью для B. orc и B. parvus - окончательного ответа пока не имеется.
Borophagus orc
ГОЛОТИП: UF 13180 (AMNH 98085), правая нижнечелюстная ветвь из хемфиллия Флориды.
ОБСУЖДЕНИЕ: Уэбб (Webb, 1969) отметил довольно смешанную примитивную и прогрессивную морфологию B. orc в сравнении с различными видами линии Osteoborus-Borophagus и пришёл к выводу, что «вероятно, предки для некоторых видов борофагов - таких, как и O. hilli и O. progressus, были уже эволюционно прогрессивны». Поскольку все знания об этом таксоне не улучшились с первоначального описания Уэбба, его филогенетическая интерпретация до сих пор имеет такую же неопределённость.
Borophagus parvus
ГОЛОТИП: FAM 75857, череп из отложений позднего хемфиллия Аризоны.
ОБСУЖДЕНИЕ: за исключением небольшого размера, B. parvus выглядит морфологически промежуточным между более прогрессивным B. secundus и примитивным B. pugnator. Поскольку B. orc в филогенетическом анализе меньше по размеру и более примитивен, возникает соблазн предположить, что B. parvus стоит в начале тенденции к увеличению размера у B. secundus и других, более прогрессивных таксонов.
Альтернативой такому сценарию выглядит то, что небольшой размер B. orc и B. parvus является результатом параллельной эволюционной линии.
Borophagus secundus 
Borophagus secundus FAM 23350
ГОЛОТИП: AMNH 13831, левая нижнечелюстая ветвь из позднехемфиллийских отложений Небраски.
ОБСУЖДЕНИЕ: Мэтью и Кук (Matthew and Cook, 1909) изначально описали его как Aelurodon saevus secundus, полагая тесно связанным с Borophagus. Чуть позже Мэтью (1924) представлял другую форму - Hyaenognathus direptor (AMNH 18919), почему-то не сравнивая его с более ранними таксонами. Материал secundus и direptor из Snake Creek ограничен фрагментарными верхними и нижними челюстями, но в тоже время демонстрируют общность дентальной морфологии, что позволяет уверенно их идентифицировать как единую форму наряду с образцами из Техаса и Канзаса.
B. secundus является самым ранним таксоном, который уже демонстрирует краниальные и дентальные пропорции прогрессивной борофагины: максимально укороченный рострум, расширенное нёбо, редуцированные премоляры и массивный, высокоронкованный p4. 
Borophagus hilli
ГОЛОТИП: TWM 1558 (AMNH 113946), правая передняя часть черепа из позднего хемфиллия Техаса.
ОБСУЖДЕНИЕ: Джонстон (Johnston, 1939) сравнивал B. hilli с В. secundus и B. diversidens, полагая, что hilli является промежуточным таксоном. Вскоре после этого Хиббард (Hibbard, 1944) описал новую форму Osteoborus progressus, отметив, что его продвинутые характеристики разделяют все признаки, но без упоминания о раннем именовании. Хотя на самом деле с учётом датировки и морфологии, O. progressus вполне гомологичен В. hilli.
Наличествуют и другие таксономические именования но, согласно Уонгу, большинство образцов из диапазона поздний хемфиллий – раний бланк (в том числе описанные, как O. crassapineatus) конспецифичны с B. hilli. Не исключено, что и образцы B. diversidens из Blancan I и II также относятся к этому таксону (по размеру и морфологии).
Borophagus dudleyi
ГОЛОТИП: MCZ 3688, череп с почти полным зубным рядом из позднего хемфиллия Флориды.
ОБСУЖДЕНИЕ: диагностика B. dudleyi затруднительна ввиду вариативности дентальной морфометрии и морфологии, хотя это действительно высокоразвитый борофаг (расширенное нёбо, удлинённый парокциптальный выступ). Уонг (Wang, 1994) поступил корректно с учётом сравнительной морфологии, позиционируя B. dudleyi как сестринскую линию к терминальному B. diversidens.
Borophagus diversidens 
ГОЛОТИП: TMM 40287-10 (AMNH 129875), левая фрагментальная нижнечелюстная ветвь из позднего бланка Техаса.
ОБСУЖДЕНИЕ: имел очень богатую таксономическую историю, так как именовался как: Felis hillianus (Cope, 1892), Hyaenognathus pachyodon (Merriam, 1903), H. dubius (Merriam, 1903), H. matthewi (Freudenberg, 1910), H. solus (Stock, 1932).
С тех пор общий консенсус, по-видимому, постепенно концентрируется на B. diversidens как на единственных и наиболее часто упоминаемых гиенообразных канид в бланке Северной Америки. Тот факт, что наличествует лишь одна форма этой собаки в большинстве местонахождений, предполагает одну непрерывную линию (анагенетическую эволюцию) в конце бланка. В качестве терминального таксона Borophaginae B. diversidens, безусловно, является наиболее специализированной формой. Морфологические тенденции в этом направлении направлены на сокращение рострума и нижних челюстей, расширение лицевой области, уменьшение премоляров в сравнении с чрезвычайно развитым P4, увеличение паракона на M1, увеличение и латеральное смещение p4, удлинение m1 и уменьшение m2.
Очевидно, что в позднем бланке (Blancan V, приблизительно 2 млн.) борофаги превратились в чрезвычайно успешную и порой доминирующую группу хищников на североамериканском континенте.
ОБЩИЙ ВЫВОД: безусловно, борофагины выглядят морфологически очень вариативными - в отличие от базальных форм, в процессе эволюции они постепенно приобретали дополнительные характеристики, которые легко отличают их как от геспероциновых, так и от собственно собачьих.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#955 30 December 2018 13:52:45
- Алекс
- Сан Саныч - Модератор

- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Ископаемые псовые (Canidae)
Игорь ,спасибо за проделанную работу ! Эпицион Хаудена -скелет . 
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#956 30 December 2018 14:12:29
- Eugene
- Модератор

- Откуда: Москва
- Зарегистрирован: 12 March 2013
- Сообщений: 5056
Re: Ископаемые псовые (Canidae)
Ёксель-моксель, что творится-то! Все, буду читать...
Спасибо, Игорь, огромное!!!
Неактивен
#957 30 December 2018 15:45:43
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Подсемейство Caninae Fischer de Waldheim, 1817
Диагностические синапоморфии включают небольшие и утоньшённые горизонтальные ветви нижней челюсти; узкие и удлинённые премоляры; задние бугорки на p2-p3 редуцированные или отсутствуют; Р2-Р3 и p2-p4 разделены короткими диастемами; P4 утоньшённый с небольшим протоконом, тригонид на m1 удлинённый (особенно параконид); парокциптальный выступ направлен вентрально и слитый с буллой - эти особенности приобретаются поэтапно в период ранней истории Caninae с узкими и удлинёнными премолярами, обозначающими начальную дивергенцию канин от борофагов.
Филогенетический анализ, включая ископаемые таксоны, демонстрирует, что большинство признаков были приобретены в ходе эволюции базальным родом Leptocyon.
LEPTOCYON Matthew, 1918
ТИПОВОЙ ВИД: Leptocyon vafer Leidy,1858.
РАСПРОСТРАНЕНИЕ: ореллий – поздний кларендоний (средний олигоцен – конец среднего миоцена) Северной Америки (30-10 млн л. н.)
ДИАГНОЗ: Leptocyon отличается от лисьих следующими примитивными характеристиками: большое посторбитальное сужение; с I1-I3 с медиальными бугорками; M1 и M2 с крупными парастилями и пр.
ОБСУЖДЕНИЕ: когда Лейди (Leidy, 1858) первоначально описывал эту форму, то указал, что она мало отличается от современной рыжей лисицы. Данное мнение присутствовало до работы Мэтью (Matthew, 1918), позиционировавшего его как переходная форма от Canis к Vulpes. Несколько видов, которые считаются принадлежащими к Leptocyon (включая L. mollis, L. vulpinus, L. gregorii) первоначально были описаны как Nothocyon. Уэбб (Webb, 1969) первым выделил морфологические функции, которые следует использовать для выделения Leptocyon в отдельный таксон.
Синапоморфии, объединяющие Leptocyon и Vulpes в первую очередь - нижнечелюстной изгиб и удлинённая область премоляров с диастемой, в то время как у геспероционов и борофагин премоляры имеют бОльшую тенденцию к редукции.
Leptocyon sp. A
МАТЕРИАЛ: UNSM 25354, левый нижнечелюстной фрагмент с p3-p4 и m1 из ореллия Небраски.
ОБСУЖДЕНИЕ: этот образец является единственным доказательством наличия Leptocyon [/i] и, следовательно, Caninae в олигоцене. Премоляры являются наиболее диагностическими элементами. Этот образец немного меньше, чем голотип и образцы L. douglassi, отличаясь отсутствием заднего выступа на p3, относительно коротким m1 с редуцированным и лабиально спроецированным параконидом (напоминая олигоценовых геспероционовых) и пр.
[i] Leptocyon mollisMerriam, 1906
ГОЛОТИП: UCMP 90 (AMNH 129683) череп с зубным рядом из раннего ариакария Небраски.
ДИАГНОЗ: отличается от L. douglassi большим размером и пропорционально более крупными М1 и М2; от L. vulpinus меньшими коронками премоляров; от L. gregorii более широким черепом и крупными P3-Р4.
Leptocyon douglassi Tedford et al., 2009
Leptocyon douglassi (CMNH 1264), фрагмент голотипа
ГОЛОТИП: CMNH 1264, передняя часть черепа и фрагмент левой нижнечелюстной ветви из раннего арикария Монтаны.
ДИАГНОЗ: отличается от всех других арикарийских видов прогрессивной морфологией p3, m2 с длинным талонидом (90% длины тригонида) и отсутствием параконида на m2.
ОБСУЖДЕНИЕ: редукция заднего бугорка на р3 является производной функцией этого раннего вида Leptocyon. P4 имеет крупный протокон и небольшой парастилярный гребень как у L. mollis и L. vulpinus, но отличаясь от тонких верхний карнассиалов у L. gregorii. M1 пропорционально меньше, чем у L. mollis, хотя морфологически с ним сходен.
Leptocyon vulpinus Matthew, 1907
Leptocyon vulpinus (AMNH 12883), фрагмент голотипа
ГОЛОТИП: AMNH 12883, левая нижнечелюстная ветвь из среднего арикария Южной Дакоты.
ДИАГНОЗ: отличается от L. douglassi и L. gregorii большим размером, выраженным парастилем на M1, пропорционально крупным m2 с большим параконидом; от L. mollis размером и высококоронкованными премолярами.
ОБСУЖДЕНИЕ: Макдональд (Macdonald, 1963) размещал этот вид в Cynodesmus согласно морфологии m1, однако другими признаками Leptocyon [/i] сильно отличается от геспероционовых. Также эта форма отличается от L. gregorii из слоёв эквивалентного возраста более широким P4 и крупным парастилем на M1.
[i] Leptocyon delicatus Loomis, 1932
ГОЛОТИП: ACM 31102, фрагмент левой нижнечелюстной ветви из среднего арикария Южной Дакоты.
ДИАГНОЗ: отличается от других видов очень небольшими размерами (примерно на 40%); премолярные и цингулумные бугорки меньше, чем у современных таксонов; метаконид на m1 пропорционально больше и выше относительно размера и высоты протоконида; на m2 отсутствует параконид.
ОБСУЖДЕНИЕ: по данному образцу Макдональд (Macdonald, 1963) описал новый род Neocynodesmus, дифференцировав от Cynodesmus, но отметил, что эта форма, возможно, эволюционировала от более старых форм.
Большинство из признаков L. delicatus аналогичны Leptocyon: морфология энтоконида на m1, узкие и удлинённые премоляры; p4 с редуцированным задним бугорком; отделённые диастемой премоляры и пр.
Leptocyon sp. B
МАТЕРИАЛ: FAM 97247, правый фрагмент нижнечелюстной ветви из позднего хемингфордия (? раннего барстовия) Небраски.
ОБСУЖДЕНИЕ: M1 выглядит более прогрессивным по отношению к L. delicatus в том, что тригонид более открыт и удлинён по отношению к длине талонида, а параконид менее заострённый. Морфологически образец несколько сходен с L. douglassi.
Leptocyon gregorii Matthew, 1907
Leptocyon gregorii (FAM 49063)
ГОЛОТИП: AMNH 12879, задняя область черепа с частью зубного ряда и фрагменты нижней челюсти из среднего арикария Южной Дакоты.
ДИАГНОЗ: отличается от других лептоционов меньшим размером и несколько примитивными дентальными характеристиками.
Leptocyon leidyi Tedford et al., 2009
Leptocyon leidyi: A–С ранний барстовий Колорадо (FAM 28345), D–F поздний барстовий Калифорнии (FAM 27535)
ГОЛОТИП: UNSM 25715, частичный череп из раннего хемингфордия Небраски
ДИАГНОЗ И ОБСУЖДЕНИЕ: отличается от других видов зубными признаками, выраженными в морфологии вторых коренных зубов. Однако дентальные особенности L. leidyi в большинстве случаев оказывается плезиоморфными по отношению к L. vafer, и в этом отношении L. leidyi можно считать предковым для поздних видов Leptocyon.
Leptocyon vafer Leidy, 1858
Leptocyon vafer (FAM 62763)
ДИАГНОЗ: отличается от L. leidyi большим размером, m2 короче m1 с увеличенным цингулумом и крупным метаконидом. Отличается от L. tejonensis более удлинённой лицевой частью черепа и более длинными премолярами, от L. matthewi - меньшим размером и морфологией коренных зубов.
ОБСУЖДЕНИЕ: впервые описан Лейди (Leidy, 1858) как небольшой вид койота. Позднее Мэтью (Matthew, 1918) признал, что морфология нижней челюсти и зубной ряд отличался как от Canis, так и от Vulpes, обозначив C. vafer как новый род, Leptocyon.
Leptocyon matthewi Tedford et al., 2009
Leptocyon matthewi (FAM 25198), голотип
ГОЛОТИП: FAM 25198, нижняя челюсть с зубным рядом из позднего кларендония Небраски.
ДИАГНОЗ И ОБСУЖДЕНИЕ: резко отличается от [i]L. vafer крупным m1 без какого-либо совпадения в размерном диапазоне. Кроме того, все диагностические признаки легко отличают L. matthewi от L. vafer и других видов Leptocyon.
Leptocyon tejonensis Tedford et al., 2009
ГОЛОТИП: LACM 16719, фрагмент правой нижнечелюстной ветви из раннего кларендония Калифорнии.
ДИАГНОЗ И ОБСУЖДЕНИЕ: прогрессивные признаки, которые отличают L. tejonensis от L. leidyi и L. vafer заключаются в укорочении лицевой части, редукции премоляров, уширении р4, талониде на m2 более 90% тригонида и пр. Премоляры, однако, заметно отличаются от всех других видов большей укороченностью и уширенностью. Этим и укрупнённостью метаконида на m2 он сходен с прогрессивными Metalopex или Urocyon, включая протостилид на m1 – коэволюционно приобретёнными характеристиками.
Триба Vulpini Hemprich and Ehrenberg, 1832
Современные лисьи и ископаемые таксоны, с ними связанные, филогенетически вполне группируются, отличаясь от других Caninae широким парокципитальным выступом, иной позицией метаконуля и постпротокристы на М2, увеличением метаконида на m2 (превышающем размер протоконида) и другими признаками.
UROCYON (серая лисица) Baird, 1857
ТИПОВОЙ ВИД: Urocyon cinereoargenteus Schreber, 1775.
РАСПРОСТРАНЕНИЕ: средний хемфиллий (поздний миоцен) – ныне
ДИАГНОЗ: отличается от других лисьих следующими апоморфиями: более расширенное сжатие на дорсальной поверхности заглазничного выступа, лобная область более уширенная, медиальные выступы на I1-I2 отсутствуют; клыки небольшие; энтоконид и гипоконид на m1 обычно связаны гребнями, гипоконулид несколько укрупнён; тригонид на m2 короче параконида и пр.
ОБСУЖДЕНИЕ: считается сестринским таксоном для Metalopex в отличие от других Vulpini - у него имеется протостилид на m1 и m2 и выраженный наружный цингулум на m2. Площадь этого зуба небольшая, хотя наличествует паракристид. У наиболее прогрессивных уроционов морфология моляров несколько отличается от ископаемых таксонов.
Urocyon webbi Tedford et al., 2009
ГОЛОТИП: UF 19407, правая нижнечелюстная ветвь из среднего хемфиллия Флориды.
ДИАГНОЗ: отличается от видов Metalopex следующими апоморфиями: P4 с хорошо развитым передним цингулумом и m2 c относительно крупным параконидом, редукцией р2 и пр.
ОБСУЖДЕНИЕ: является примитивней по отношению к другим видам, но очень схож с таковыми в форме премоляров. Нижняя челюсть U. webbi крупнее и массивнее, чем у других Urocyon.
Urocyon progressus Stevens, 1965
ГОЛОТИП: UMMP 37157, левая теменная кость из раннего бланка Канзаса.
ОБСУЖДЕНИЕ: диагноз Стивенса (Stevens, 1965) может быть сомнителен ввиду отсутствия других образцов кроме типового. Впрочем, Бьорк (Bjork, 1973) упоминал о частичном гнатокраниальном материале, возможно относимом к U. progressus. Однако наличествуют отличия в морфологии теменной кости.
Urocyon galushai Tedford et al., 2009
ГОЛОТИП: FAM 63104, правая и левая фрагментарные нижнечелюстные ветви из позднего бланка Аризоны.
ДИАГНОЗ: отличается от U. citrinus большими премолярами; меньшим m2 с более короткими тригонидом и талонидом, от U. minicephalus и U. cineroargenteus более крупными, высококоронованными премолярами и удлинённым m2.
ОБСУЖДЕНИЕ: Гетц (Getz, 1960) описывал U. atwaterensis, основываясь на отличительной морфологии M2, сходной с серией образцов современного U. cinereoargenteus. В тоже время степень развития метаконуля этого зуба у уроционов достаточно вариативна и при сравнении с U. minicephalus и U. cinereoargenteus он выглядит примитивней. С другой стороны, U. galushai уникален в удлинённости горизонтальной нижнечелюстной ветви наряду с большими передними премолярами и прочими признаками, которые позволяют выделить эту форму в отдельный таксон видового ранга.
Urocyon citrinus Tedford et al., 2009
ГОЛОТИП: UF 18060, фрагментарные связанные нижнечелюстные ветви из раннего ирвингтония Флориды.
ДИАГНОЗ: аутапоморфии Тедфордом (Tedford et al., 2009) не распознаны, но данный вид морфологически принадлежит к этому таксону, отличаясь от других форм некоторыми плезиоморфными признаками.
ОБСУЖДЕНИЕ: схож с U. webbi, но это сходство скорее, основано на примитивных характеристиках группы. Диагноз акцентируется на прогрессивных морфологических признаках, которые U. citrinus разделяет с другими видами Urocyon.
Urocyon minicephalus Martin, 1974
Urocyon minicephalus (UF 13146), голотип
ГОЛОТИП: UF 13146 почти полный череп из позднего ирвингтония Флориды.
ДИАГНОЗ: отличается от U. citrinus следующими апоморфиями: менее удлинённым m1, относительно меньшим m2 с коротким тригонидом и отсутствующим параконидом. Отличается от U. galushai меньшими и низкокоронкованными премолярами; укороченным тригонидом на m2; от U. cinereoargenteus меньшим черепом, более заметным посторбитальным сужением, M1 с низкокоронованными параконом и метаконом, редуцированным протостилидом на m1; отличается от U. c. scotti меньшими размерами с относительно широкой мордой, большем посторбитальном сужении, узким затылком и пр.
ОБСУЖДЕНИЕ: Мартин (Martin, 1974) описал череп U. minicephalus как отличающийся от современной серой лисицы краниальными признаками и серией дентальных – в целом, они выглядят несколько примитивнее, чем у современных.
METALOPEX Tedford and Wang, 2008
Гипокарниворный Metalopex и его сестринский таксон Urocyon образуют кладу, объединённую удлинённым талонидом на m2, высокоронкованными премолярами, протостилидом на m1-m2, более квадратной формой верхних коренных зубов и своеобразной изоляцией p2 удлинёнными диастемами. Возможно, данные признаки несколько объединяют эти таксоны с Protocyon и Otocyon.
ТИПОВАЯ ФОРМА: Metalopex merriami Tedford and Wang, 2008.
РАСПРОСТРАНЕНИЕ: поздний кларендоний – ранний хемфиллий (поздний миоцен) Северной Америки (11-9 млн л.н.)
ДИАГНОЗ: прогрессивные признаки, которые отличают Metalopex от Vulpes: крупный мастоидный отросток, кубоидная форма коренных зубов и другие дентальные характеристики.
ОБСУЖДЕНИЕ: главным образом отличается от Vulpes и Urocyon в уникальной морфологии М2 с отсутствующей постпротокристой, но наличествующим метаконусом; а также в сохранении примитивной формы верхних резцов с медиальным бугорком на I3, в относительно прогрессивном расширении мастоидного отростка (в сравнении с Leptocyon), увеличенном m3.
Metalopex macconnelli Tedford et al., 2009
Metalopex macconnelli (LACM 55237), голотип
ГОЛОТИП: LACM 55237, череп с зубным рядом из позднего кларендония Калифорнии.
ДИАГНОЗ: отличается от M. merriami меньшим размером, позицией височных и затылочных гребней, крупной буллой, боковыми выступами на резцах, формой коренных зубов.
ОБСУЖДЕНИЕ: морфологические особенности по большей части напоминают Leptocyon, но относительное расширение мастоидного отростка подобно голотипу. Аналогично, мандибула является удлинённой и тонкой, весьма отличаясь морфологией восходящей ветви. Хотя некоторые образцы (к примеру, NMNH 23883), приписываемые этому таксону, отличаются от типового образца M. macconnelli, Тедфорд (Tedford et al., 2009) считает вариативность в морфологии р4-m3 признаками, демонстрирующими хроноклинальные отличия в размерах и морфологических вариациях (популяционные, половые, возрастные и индивидуальные).
Metalopex merriami Tedford and Wang, 2008
Metalopex merriami (FAM49282), голотип
ГОЛОТИП: FAM 49282, частичный череп с зубным рядом из раннего хемфиллия Орегона.
ДИАГНОЗ: отличается от M. macconnelli более крупным размером, расширенной лицевой областью, топологией затылочной, расширенными буллами, боковыми бугорками на резцах, иной морфологией бугорков на коренных зубах.
ОБСУЖДЕНИЕ: существуют значительные различия в размерах и морфологии с M. macconnelli из разных местонахождений. За исключением одного образца из Black Hawk Ranch, который демонстрирует явные морфологические аспекты Urocyon, имеющийся ископаемый материал из разных локаций указывает на значительную размерную вариативность и морфологическую интерградацию, предполагая анагенетические изменения от позднего кларендония до раннего хемфиллия для данного таксона.
Metalopex bakeri Tedford et al., 2009
ГОЛОТИП: FAM 62970, левая фрагментарная нижнечелюстная ветвь из среднего хемфиллия Техаса.
ДИАГНОЗ: отличается от M. merriami более короткими премолярами и укороченным премолярным рядом без выраженных диастем (кроме отделяющих p2); укороченный тригонид на m1 и пр.
ОБСУЖДЕНИЕ: зубной ряд M. bakeri отличается многочисленными аутапоморфиями. Премоляры заметно короче и более высокоронкованы, чем у М. merriami; P2 отделён диастемой, которая короче, чем у M. merriami; M1 очень массивен - ширина талонида равна или пропорционально больше в сравнении с тригонидом, чем у М. merriami и прочие признаки. Также в черепе налицо характеристики, формирующие мнение о более крупном и удлинённом лобном сжатии, чем у большинства вульпин, кроме того налицо явная разница в топологии сагиттального и затылочного гребней.
PROTOTOCYON Pohle, 1928
ТИПОВОЙ ВИД: Prototocyon recki Pohle, 1928
РАСПРОСТРАНЕНИЕ: поздний плиоцен – ранний плейстоцен (2.0–1.7 млн) Танзании
ДИАГНОЗ: небольшая лисица, близкая к Otocyon; лобные пазухи и сагиттальный гребень отсутствуют; премоляры небольшие; хищнические зубы с редуцированным парастилем, моляры относительно крупные.
ОБСУЖДЕНИЕ: известен на основе материала нескольких особей из Olduvai, Главным отличием от Otocyon являются более примитивные дентальные характеристики. Остатки сходных псовых также регистрируются в Laetoli, но принадлежат ли они Prototocyon, ещё предстоит выяснить.
OTOCYON (большеухая лисица) Müller, 1835
РАСПРОСТРАНЕНИЕ: 300 тлн (ср. плейстоцен) – ныне, Африка
ДИАГНОЗ: мелкоразмерные лисицы с гипертрофированным числом коренных зубов, премоляры небольшие; P4 и m1 гипокарнировидные; m1 с редуцированным тригонидом и удлинённым талонидом; P4 с очень коротким метастилем.
ТИПОВОЙ ВИД: O. megalotis Desmarest, 1822
ОБСУЖДЕНИЕ: ископаемые образцы O. megalotis известны только из местонахождения Лайньямок (Lainyamok) Кении.
VULPES (обыкновенная лисица) Frisch, 1775
Vulpes stenognathus (FAM 49284), поздний хемфиллий Невады
ДИАГНОЗ: Vulpes отличается от Leptocyon следующими синапоморфиями, объединяющими их с высшими Caninae: отсутствие боковых бугорков на I3; скуловая кость сильно изогнута; парастиль на M1 очень редуцирован; метакон на M2 наличествует; задние конечности по отношению к передним удлинены и пр.
РАСПРОСТРАНЕНИЕ: 7 млн (поздний миоцен) – ныне, Палеарктика.
ИСКОПАЕМЫЕ ВИДЫ: Vulpes riffautae de Bonis et al., 2007; V. skinneri Adam Hartstone-Rose et al., 2013; V. kernensis Tedford et al., 2009; V. velox Say, 1823; V. stenognathus Savage, 1941; V. rueppelli Schinz, 1825; V. hassani Geraads, 2011; V. praeglacialis Kormos, 1932
ОБСУЖДЕНИЕ: линия вульпин весьма богата разнообразием, демонстрируя высокую адаптивность в процессе эволюции. Древнейшие известные ископаемые Vulpes известны как V. riffautae из позднего миоцена Чада (Toros-Menallа) – самая ранняя фиксация лисьих в Старом Свете, чьи морфологические характеристики отличают этот образец от всех остальных лисиц. Наличие Vulpes и Eucyon в Африке, а также на юго - западе Европы в конце миоцена может свидетельствовать о том, что эти псовые мигрировали в Европу из Африки через транссредиземноморской маршрут. Гипокарниворными характеристиками V. riffautae очень отличается как от современных лисиц, так и от феннеков. Другая африканская лиса – V. skinneri из раннего плейстоцена Южной Африки. В статье, описывающей образцы, авторы утверждают, что «экземпляры достаточно отличаются, чтобы относиться к новому виду». В то же время она моложе V. riffautae на 5 млн лет и единственным, несколько связующим звеном, является V. hassani из миоцен-плиоцена в северо-западной Африке. Это тоже энигматичная форма – морфологически она мало чем отличается от современной серой лисы V. rueppelli, однако большинство образцов метрически крупнее нынешней формы и это не позволяет объединить их в один таксон.
Из Европы известна V. praeglacialis – эндемичная форма из раннего плейстоцена Греции. В Северной Америке – мелкоразмерный позднемиоценовый (хемфиллий) V. kernensis. Плиоценовый V. stenognathus широко распространён по всему материку от тихоокеанского побережья до Флориды в бланкских местонахождениях, хотя встречается редко. V. kernensis известен только из раннего хемфиллия Калифорнии, отличался от сосуществовавшего V. stenognathus длиной зубного ряда, укорочением премоляров и пр. Раннеирвингтонский V. velox выглядит единственным представителем лисиц с раннего по средний плейстоцен Северной Америки, так как V. vulpes и V. lagopus появляются только в его конце.
ПРИМЕЧАНИЯ: Фенек V. zerda иногда причисляется к особому роду Fennecus (Fennecus zerda Zimmermann, 1780).
(?) ALOPEX (песец) Linnaeus, 1758
СИНОНИМИЧЕСКИЕ ЗНАЧЕНИЯ: Vulpes lagopus
РАСПРОСТРАНЕНИЕ: поздний плейстоцен циркумарктического региона – ныне
ДИАГНОЗ: характерно увеличение режущих частей нижнего и верхнего хищнических зубов и удлинение моляров. В верхнем зубном ряду функциональные особенности маскируют корреляции, возникающие в результате общей истории развития морфологических структур. Дентальная морфология между континентальными и островными формами вариативна
ОБСУЖДЕНИЕ: ранее определялся в монотипичный род Alopex, но последние генетические данные помещают его в род Vulpes. В то же время до сих пор не имеется консенсусного мнения о генезисе этой формы. Ввиду находки Vulpes qiuzhudingi (Wang et al. , 2014) из позднего плиоцена Тибета с выраженной гиперкарниворностью хищнических зубов имеется гипотеза о происхождении песцов именно из этой области.
ВЫВОД: дентальная морфовариативность канид «лисьей» группы стимулирована высокой конкурентностью с другими хищническими таксонами, с диапазоном от омнио- и инсектофагии в южных и средних широтах к карнофагии в высоких. Пример – феннек: большие отличия в краниуме ввиду расширенных булл и сопутствующие изменения, резко отличающие его от других вульпин. Или песец - архетипически он, конечно, тоже - лисица, но зубки-то чуть другие. И подчас даже меньшими отличиями руководствуются исследователи, отделяя один род псовых от другого.
К слову, аналогичная картина наблюдается у медведей – краниодентальная морфология белого медведя резко отличается от остальных урсид.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#958 30 December 2018 15:54:10
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Триба Canini Fischer de Waldheim, 1817
Объединяет современных - собак, волков, койотов, шакалов и южноамериканских псовых, морфологически характеризуясь как сестринская группа к трибе Vulpini. Предполагается монофилетическое происхождение (Tedford et al., 1995) с дивергенцией ‘‘South American foxes’’ (Wayne et al., 1997) – Cerdocyonina, филетически связанных в комбинированном анализе морфологических и молекулярных признаков. Очевидно, что фоссильные виды, близкие к современным Cerdocyon, Chrysocyon и, возможно, Theriodictis, сформировались в северных широтах задолго до 3 млн. лет - до возникновения Панамского перешейка, что объясняет изоляцию рода Nyctereutes.
Филогенетический анализ показывает, что виды Eucyon являются сестринскими ко всем другим видам Canini вместо того, чтобы лежать у основания Canina. Ареал этой формы в миоцене ограничивался Северной Америкой, появившись в плиоцене и в Старом Свете.
EUCYON Tedford and Qiu, 1996
РАСПРОСТРАНЕНИЕ: поздний миоцен – ранний миоцен Северной Америки, Азии, Европы и Африки
ДИАГНОЗ: отличается от Vulpini синапоморфиями, которыми обладают все остальные представители Canini: гипертрофия лобной пазухи, расширенный кзади парокципитальный выступ с харктерным кончиком, увеличенный мастоидный отросток. В то же время у Eucyon отсутствуют некоторые характеристики, присущие Canini – в основном, касающиеся дентальной морфологии.
СЕВЕРНАЯ АМЕРИКА:
Eucyon davisi Merriam, 1911
РАСПРОСТРАНЕНИЕ: хемфиллий (поздний миоцен – ранний плиоцен Северной Америки
ДИАГНОЗ: отличается от E. skinneri большим размером; более длинными челюстями, высококоронованными премолярами; относительно удлинённым тригонидом на m1; от Canis ferox и C. lepophagus меньшими размерами и более робустным черепом, менее выраженными скулами; морфологией лобного синуса, сагиттального гребня и щечных зубов.
ОБСУЖДЕНИЕ: наличествует много гнатодентальных образцов, относящихся к E. davisi, но полные черепа, представляющие североамериканские популяции, пока неизвестны. Краниальные описания, в основном, основаны на трёх фрагментарных образцах FAM 63010, 63007 и 63009. Финальная диагностика может быть основана только при наличии сравнительного материала.
Eucyon skinneri Tedford et al., 2009
ГОЛОТИП: FAM 25143, нижнечелюстной фрагмент с зубным рядом из позднего кларендония Небраски.
ДИАГНОЗ: синапоморфии, которые объединяют E. skinneri с E. davisi[/i] и другими канинами: m1 с хорошо развитым энтоконидом и небольшим гипоконулидом. Примитивность заключается в меньшем размере, укороченной челюсти со сниженным венечным отростком а также морфологии щечных зубов.
ОБСУЖДЕНИЕ: сравнение с Leptocyon и E. davisi демонстрирует, что нижняя челюсть и зубной ряд крупнее и массивнее. Кроме того, талонид на m1 у E. skinneri пропорционально шире и с более развитым энтоконидом, а также отсутствует поперечный гребень между энтоконидом и гипоконидом.
АЗИЯ:
Eucyon sp. А
МЕСТОНАХОЖДЕНИЕ: Хиргис Нур (Khirgis Nur), Монголия.
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: поздний миоцен.
ОБСУЖДЕНИЕ: ранее упоминание фиксация рода в Китае из верхнего баодэ (поздний миоцен) из бассейна Юше в данный момент является недействительным (Wang, 2006). Новые данные из Монголии показывают, что самое раннее появление рода в Азии совпадает с фиксацией в Европе и Африке, подтверждая, что миграция Eucyon из Америки в Старый Свет было синхронным.
Eucyon davisi (?) Merriam, 1911
ГЕОГРАФИЧЕСКИЙ ДИАПАЗОН В АЗИИ: Китай, Монголия, Казахстан.
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: поздний миоцен – ранний плиоцен
ОБСУЖДЕНИЕ: не отличается на определенном уровне от типичных североамериканскихе видов E. davisi. В дополнение к материалу из Юше имеются образцы, относящиеся к E. davisi из раннего плиоцена Монголии (Khirgis Nur) и Казахстана (Павлодар) (Sotnikova & Rook 2009).
Eucyon zhoui Tedford & Qiu, 1996
МЕСТОНАХОЖДЕНИЕ: Юше (Yushe Basin), Китай.
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: ранний – средний плиоцен.
ОБСУЖДЕНИЕ: крупнее E. davisi и отличается от него своеобразными краниальными пропорциями, одновременно демонстрируя больше прогрессивных черт как в нижней челюсти, так и в морфологии зубов. Согласно Тедфорду и Цю (Tedford and Qiu, 1996), может быть ближе к группе Canis, чем к E. davisi
Eucyon minor Teilhard de Chardin & Piveteau, 1930
МЕСТОНАХОЖДЕНИЕ: Нихэвань (Nihewan), Китай.
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: поздний плиоцен Китая, Монголии и Забайкалья.
ОБСУЖДЕНИЕ: образцы описываются как койотоподобные собаки, демонстрируя сходную морфологию с C. lepophagus, но меньшую размерность.
Eucyon marinae Spassov & Rook, 2007
МЕСТОНАХОЖДЕНИЕ: Мухор-Эриг (Muhor-Erig), Монголия.
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: поздний плиоцен.
ОБСУЖДЕНИЕ: является относительно крупным видом, сопоставимым по размеру с E. davisi и E. monticinensis, отличающимся грациальной мандибулой с удлинённым рострумом и очень длинным премолярным рядом.
ЕВРОПА:
Eucyon cipio Crusafont Pairo, 1950
МЕСТОНАХОЖДЕНИЕ: Консуд (Concud), Испания.
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: туролий (MN-12), поздний миоцен
ОБСУЖДЕНИЕ: представлен типовым образцом (правая верхнечелюстная кость с P3-M2) и изолированным правым нижним карнассиалом (m1). Морфологически выглядит примитивным, наводя на мысль о ранней стадии эволюции перед дифференциацией рода Canis.
Eucyon monticinensis Rook, 1992
МЕСТОНАХОЖДЕНИЕ: гипсовый карьер (Brisighella), Италия.
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: туролий (MN-13), поздний миоцен
ОБСУЖДЕНИЕ: мелкоразмерная форма; вместе с E. davisi и E. intrepidus один из ранних представителей рода. Возможно, к нему относится также материал из позднемиоценового испанского местонахождения Venta del Moro, ранее описанный как Nyctereutes (Morales & Aguirre 1976; Моралес, 1984).
Eucyon odessanus Odintzov, 1967
МЕСТОНАХОЖДЕНИЕ: Одесские катакомбы (Украина)
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: ранний плиоцен (русциний, MN 15) Молдовы, Украины, Греции и Турции.
ОБСУЖДЕНИЕ: крупный образец из одесских катакомб описывается как Vulpes odessana, хотя скорее всего, принадлежит роду Eucyon. Зубной ряд E. odessanus размерно аналогичен E. davisi, но демонстрирует некоторые различия в премолярах.
Eucyon adoxus (Martin, 1973)
МЕСТОНАХОЖДЕНИЕ: Сан-Эстев (St-Esteve), Франция
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: ранний плиоцен (русциний, MN 15) Франции и Испании.
ОБСУЖДЕНИЕ: характеризуется удлинённой нижней челюстью и, очевидно, не является лисоподобным псовым, как утверждалось некоторыми авторами (Pons Moya & Crusafont Pairo 1978). Хотя, с другой стороны, некоторые из прогрессивных признаков, являющимися синапоморфиями для канин, могут представлять таксон, отличающийся от Eucyon.
Eucyon debonisi Morales & Aguirre 1976:
МЕСТОНАХОЖДЕНИЕ: Вента-дель-Моро, Испания
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: поздний миоцен (туролий, MN 13)
ОБСУЖДЕНИЕ: имеет меньший зубной ряд, чем E. davisi за исключением M2. Также имеются отличия в морфологии щечных зубов. В то же время наличествует некоторое сходство с E. monticinensis.
«Canis» kuruksaensis Sotnikova, 1989
ГОЛОТИП: ПИН РАН М 3120-251, осевой череп с зубным рядом.
МЕСТОНАХОЖДЕНИЕ: Таджикистан, Куруксай, точка Наврухо
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: поздний плиоцен (средний виллафранк).
ДИАГНОЗ: мелкий представитель, размеры черепа в пределах изменчивости современных шакалов. Лицевая часть черепа немного больше мозговой коробки. Бугорок у переднего наружного края паракона на М1 хорошо развит.
ОБСУЖДЕНИЕ: отличается от большинства эуционов более крупными зубами, однако сохраняет примитивные признаки, присущие Eucyon. Основываясь на морфологии верхних зубов, ''C.'' kuruksaensis более прогрессивен и ближе к Canis, чем все евразийские эуциономорфные псовые. По этой причине именование ''C.'' kuruksaensis даётся в кавычках.
АФРИКА:
Eucyon intrepidus Morales, Pikford & Soria, 2005
МЕСТОНАХОЖДЕНИЕ: Туген-Хиллз, Кения
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: поздний миоцен
ОБСУЖДЕНИЕ: самый ранний известный представитель Canini в Африке. Североамериканские и европейские виды (E. davisi, E. monticinensis) имеют меньшие размеры и более высокие тригониды. Также африканские виды характеризуются более коротким талонидом в сравнении с европейскими и североамериканскими Eucyon.
Eucyon wokari Garcia, 2008
МЕСТОНАХОЖДЕНИЕ: Aramis, (Эфиопия).
ХРОНОЛОГИЧЕСКИЙ ДИАПАЗОН: ранний плиоцен
ОБСУЖДЕНИЕ: размерно сопоставимый с E. intrepidus, в детальной морфологии нижних коренных зубов E. wokari резко отличается в структуре талонидов как более прогрессивными. Возможно, несколько сходен с китайским E. zhoui и, в итоге, более тесно связан с группой Canis, чем с Eucyon.
ВЫВОДЫ: род Eucyon был создан Tedford & Qiu (1996) на основании обильного ископаемого материала ранних канин с позднего миоцена Северной Америки до плиоцена Евразии. Данная форма объединяет все краниальные синапоморфии, отличающие их от Vulpini. В общем, морфология Eucyon сходна с Vulpes, поскольку они имеют множество примитивных особенностей. Тем не менее, Eucyon, как и Cerdocyonina, в сравнении с Canis, обладает небольшой лобной пазухой, в то же время разделяя с таковыми основные синапоморфные признаки.
Берта (Berta, 1988) рассматривала E. davisi в качестве базального таксона как для южноамериканских Cerdocyonina, так и для продвинутых форм. Самый ранний и примитивный E. davisi из раннего плиоцена Азии, очевидно, тесно связан с позднетуролийским E. monticinensis. Также хорошо сохранившийся краниальный материал E. davisi известен из разных плиоценовых местонахождений Китая. Другие плиоценовые евразийские эуционы - Е. odessana и E. zhoui вполне демонстрируют прогрессивные особенности, сближающие их с Canis.
Присутствие Canidae в позднем миоцене Евразии долгое время оставалась крайней редкостью, кроме С. cipio (ранее классифицируемом как Nyctereutes aff. donnezani) до находок Е. monticinensis, E. debonisi и пр. Имеется гипотеза о связи E. debonisi с африканскими формами в конце миоцена – тем более, при обильном наличии евразийских иммигрантов в Африке. Впрочем, для этого нужно больше фаунистических доказательств. Что же касается африканских видов, то радикальные сравнения всё ещё ограничены нехваткой ископаемого материала.
Прогрессивные характеристики представителей Canini среднего и позднего плиоцена предполагают, что эти псовые образуют отдельную группу эуционоподобных видов, более продвинутых в направлении Canis. К таковым формам относятся E. marinae, E. minor и ''Canis'' kuruksaensis, хотя общую атрибуцию ещё трудно обсуждать без данных о краниальной морфологии. Формально они соответствуют диагнозу Eucyon, хотя отличаются более коротким премолярным рядом и более продвинутым p4 с уменьшенным цингулумом позади второго дополнительного бугорка. Таким образом, не исключено, что целая группа Canini среднего и позднего плиоцена принадлежит к роду, отличающемуся от Canis и Eucyon.
Также, при разделении Canini на две подтрибы: южноамериканских псовых (включая Nyctereutes), и канисоморфных видов (Canis, Xenocyon, Lycaon, Cuon и др), целесообразно эуционоподобные формы неогена также выделить в отдельную таксономическую группу.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#959 30 December 2018 16:18:03
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Подтриба Cerdocyonina
Эти таксоны могут быть диагностированы следующими синапоморфиями: угловой выступ нижней челюсти может быть лишён крючкообразного конца; задний бугорок р3 редуцирован или отсутствует; иная морфология гипоконида и энтоконида на m1; а также отсутствие второго заднего дополнительного бугорка между цингулумом и первым бугорком, что объединяет Cerdocyonina с Eucyon и Canina.
CERDOCYON (саванновая лисица) Smith, 1839
ТИПОВОЙ ВИД: Cerdocyon thous Linnaeus, 1766 (современный)
РАСПРОСТРАНЕНИЕ: ранний плиоцен (6 млн) – ныне, Новый Свет
ДИАГНОЗ: синапоморфии, объединяющие Cerdocyon с Atelocynus и Speothos и отличающие его от более плезиоморфных южноамериканских канин (Dusicyon, Pseudalopex, Lycalopex) включают расширение углового выступа нижней челюсти, укороченные клыки небольшого диаметра и пр. Cerdocyon отличается от большинства видов подтрибы в том, что его нёбо не продлено дальше зубного ряда. То же самое относится к ископаемым видам Nyctereutes.
ОБСУЖДЕНИЕ: C. texanus и C.(?) avius обладают прогрессивными дентогнатальными признаками, отличающими их от других таксонов – изогнутыми кзади клыками, моляры с мезоконидом, крупный коренной зуб в пропорции к Р4 – все эти признаки очень схожи с раннеплиоценовыми Nyctereutes.
Cerdocyon texanus Tedford et al., 2009
Вверху: C. texanus (FAM 62985), голотип; внизу – C. thous (AMNH(M) 14636), современный образец из Колумбии
ГОЛОТИП: FAM 62985, левая частичная нижнечелюстная ветвь с зубным рядом из позднего хемфиллия Техаса.
РАСПРОСТРАНЕНИЕ: ранний плиоцен Техаса и Нью-Мексико.
ДИАГНОЗ: аутопоморфии, отличающие C. texanus от C. thous: I3 более сжатый; P1-P3 удлинённый и более высокоронкован; P4 удлинён с увеличенным протоконом; p1 крупный и двукоренной; тригонид на m1 длиннее карнассиала и пр.
ОБСУЖДЕНИЕ: из гнатодентальных сравнений очевидно, что C. texanus напоминает C. thous и раннеплиоценовые евразийские виды Nyctereutes - N. donnezani и N. tingi. Наличествует много специфических особенностей C. texanus, но суммарно они находятся в морфологической орбите Cerdocyon.
Этот таксон отнесён к Cerdocyon, но вполне может представлять собой представителя группы в кладогенезе Cerdocyon и сестринского таксона Nyctereutes.
Cerdocyon (?) avius Torres and Ferrusquıa, 1981
ГОЛОТИП: IGM 2903,фрагментарная нижнечелюстная ветвь без вертикальной ветви с зубным рядом и посткраниальный материал из раннего бланка Калифорнии.
ДИАГНОЗ: отличается от C. texanus наличием p1; меньшим размером зубного ряда; m1 с редуцированным метаконидом и пр.
ОБСУЖДЕНИЕ: хотя описание и сравнения этого таксона содержатся в тезисах Торреса (Torres, 1980), в резюме отмечено, что ряд гнатодентальных особенностей, несмотря на явные морфологические признаки Cerdocyonina, демонстрируют метрические пропорции Canini.
ATELOCYNUS (малая лисица) Sclater, 1883
ВКЛЮЧЁННЫЕ ПОДВИДЫ: [i]A. microtis microtis, A. m. sclateri
ОБСУЖДЕНИЕ: эндемик тропических лесов амазонского бассейна. Очевидно, что предков Atelocynus следует искать среди раннеплиоценовых псовых, мигрировавших из Северной Америки в рамках Великого американского обмена. Несмотря на явные адаптиационные особенности приспособленности к обитанию в лесных биотопах, морфологически с кустарниковой собакой, похоже, не связана.
(?) PSEUDALOPEX (южноамериканская лисица) Gray, 1837
ТИПОВОЙ ВИД: Pseudalopex griseus Gray, 1837
РАСПРОСТРАНЕНИЕ: южные районы Южной Америки
ОБСУЖДЕНИЕ: таксономия и систематика южноамериканских лисиц до сих пор дискутивна. Официально признаны три вида - андская лисица Р. culpaeus, номинативная P. griseus и парагвайская P. gymnocercus. Однако в последнее время имеется мнение, что последние две формы представляют собой лишь географические вариации отдельного рода и должны быть объединены в Lycalopex gymnocercus.
LYCALOPEX Burmeister, 1856
СОСТАВ РОДА:
Lycalopex vetulus Бразильская лисица Lund, 1842
Lycalopex sechurae Секуранская лисица Thomas, 1900
Lycalopex fulvipes Дарвиновская лисица Martin, 1837
ОБСУЖДЕНИЕ: мнение о таксономии до сих пор дискуссионно - одни считают их вполне валидным таксоном, другие до сих пор считают Pseudalopex и Lycalopex синонимичными, причём морфологически сходными с фолклендской собакой (Dusicyon australis). Скорее всего, эволюционировали примерно в конце плиоцена (2,5 млн лет назад), сохранив гипокарниворный морфотип даже в реликтовых или эндемичных формах.
THERIODICTIS Mercerat, 1891
РАСПРОСТРАНЕНИЕ: плиоцен – поздний плейстоцен Неарктики
ДИАГНОЗ: гипоконы на М1- M2 редуцированы; M2 относительно M1 небольшой с уменьшенным метаконом; венечный отросток удлинён; угловой отросток расширен дорсовентрально; метаконид на m1 отсутствует, но сохранён небольшой энтоконид; м2 относительно m1 небольшой.
СОСТАВ РОДА:
Theriodictis floridanus Tedford et al., 2009 – бланк-ирвингтоний (ранний – средний плейстоцен)
Theriodictis platensis Mercerat, 1891 – энсенадий-луханий (ср-поздний плейстоцен)
Theriodictis tarijensis Ameghino, 1902 – энсенадий-луханий (ср-поздний плейстоцен)
Theriodictis floridanus (UF 19324), голотип
ОБСУЖДЕНИЕ: образцы Theriodictis демонстрируют ряд диагностических особенностей Cerdocyonina, сближающих их с Protocyon и Dusicyon. Североамериканский T. floridanus отнесён к этому таксону на основе сходства коренных зубов и нижней челюсти в сравнении с плейстоценовыми боливийскими и аргентинскими видами, но метаконид на m1 у него ещё присутствует.
SPEOTHOS (кустарниковая собака) Lund, 1839
РАСПРОСТРАНЕНИЕ: поздний плейстоцен – ныне
СОСТАВ РОДА: Speothos pacivorus Lund, 1839 – вымерший, S. venaticus – современный
ДИАГНОЗ: череп короткий с широкими пропорциями, сагиттальный гребень низкий; височные гребни хорошо развиты. Постборбитальное сужение небольшое, носовая область укорочена, слуховые капсулы расширены. S. pacivorus отличается от S. venaticus большим размером, хорошо развитыми метаконом и гипоконом на M1, наличием M2.
Speothos pacivorus UZM L"406"180
ОБСУЖДЕНИЕ: первоначально Лунд (Lund, 1839) описывал S. pacivorus как Icticyon, отличающийся от Speothos наличием M2 и лишь позже синонимизирован Краглиевичем (Kraglievich, 1930) наряду с Abathmodon в один таксон. Биогеографическая история Speothos, похоже, полностью связана с Южной Америкой и находки S. pacivorus предполагают Бразилию как зону происхождения этой группы. Впоследствии во время позднего плейстоцена Speothos дифференцировался на два вида.
CHRYSOCYON (гривистый волк) Smith, 1839
РАСПРОСТРАНЕНИЕ: плиоцен – ныне Неарктики
ДИАГНОЗ: разделяет синапоморфии с остальными представителями Cerdocyonina, включая широкие скуловые арки, морфологию короноидного выступа, M1-M2 квадратной формы в окклюзии и пр.
СОСТАВ РОДА:
Chrysocyon nearcticus Tedford et al., 2009 – бланк (ранний плиоцен)
Chrysocyon brachyurus Illiger, 1815 – энсенадий (ср. плейстоцен) - ныне
C–D. Chrysocyon nearcticus (UA 12610), голотип; E–F. Chrysocyon brachyurus AMNH(M) 36962
ОБСУЖДЕНИЕ: открытие ископаемых форм, близко напоминающих современные виды в раннем плиоцене ясно указывает на степень филетической дифференциации Canini в то время. К примеру, диагностирование C. nearcticus опирается только на гнатодентальную морфологию и, следовательно, в диагнозе отсутствуют многие черепные и посткраниальные символы, признанные для рода. В определённой степени гнатодентальные особенности представляют собой ряд коррелированных характеристик, общих для долихоцефальных канид (Canis или Vulpes). Тем не менее, соответствие в морфологических деталях не представляет особых сомнений в таксономической принадлежности ископаемого материала.
DUSICYON Smith, 1839

РАСПРОСТРАНЕНИЕ: поздний плиоцен - ранний плейстоцен Южной Америки
ДИАГНОЗ: затылок более округлой формы, чем у Canis, скуловые кости расширены, лобные пазухи увеличены, зубы сравнительно массивны (премоляры имеют тенденцию к редукции), клыки несколько массивны. M1 с выраженным цингулумом и укрупнённым параконом в то время, как m1 имеет более остроконечный тригонид и редуцированные метаконид с энтоконидом.
A: Dusicyon avus (CEHA 5131). B: D. australis
СОСТАВ РОДА:
Dusicyon cultridens Molina, 1782 – среднеплиоценовая форма Аргентины
Dusicyon avus Burmeister, 1866 – типовой позднеплейстоценовый вид
Dusicyon sechurae Thomas, 1900 - позднеплейстоценовый вид из Эквадора
Dusicyon australis Kerr, 1792 - Фолклендская лисица
NYCTEREUTES (енотовидная собака) Temminck, 1839
Nyctereutes terblanchei
ТИПОВОЙ ВИД: Nyctereutes procyonoides Gray, 1834.
ДИАГНОЗ: лисоподобные каниды с укороченными конечностями, лицевая область узкая с низкой лобной частью, теменные бугорки с небольшим гребнем, нижняя челюсть с выраженным округлым субтреугольным выступом на заднем крае, карнассиальная область небольшая, энтоконид и гипоконид на m1 почти одинакового размера, а моляры относительно увеличены (m2 и m3 очень крупные).
РАСПРОСТРАНЕНИЕ: 9 (?) млн – ныне, Палеарктика
Nyctereutes procyonoides (AMNH 249766)
ИСКОПАЕМЫЕ ВИДЫ: N. abdeslami Geerards, 1997; N. donnezani Depéret, 1890; N. megamastoides Pomel, 1842; N. sinensis Brass, 1904; N. tingii Tedford & Qiu, 1991; N. terblanchei Ficcarelli et al., 1984; N. vinetorum Bate, 1937; N. lockwoodi Geraads et al., 2010
ОБСУЖДЕНИЕ: Остатки N. donnezani, предковой формы, были обнаружены в позднеплиоценовых местонахождениях Западной Европы (Италия, Франция, Венгрия и Румыния). Более крупная форма N. megamastoides фиксируется в том же районе в раннем плейстоцене и, очевидно, была конспецифична средне-плиоценовой . sinensis из Китая (прохорез в Азию ?).
Nyctereutes появился в Евразии в плиоцене и широко распространился в Палеарктическом регионе. Плиоценовые енотовидные собаки представлены N. donnezani и N. megamastoides в Европе, а также N. tingii и N. sinensis в Азии. Среди плиоценовых форм примитивные N. donnezani и N. tingii схожи с более прогрессивными N. megamastoides и N. sinensis в структуре и пропорциях черепа, хотя у N. donnezani и N. tingii значительно менее выраженный угловой отросток нижней челюсти и относительно большие и более четырёхугольные моляры. По сравнению с современными представителями ископаемые формы имели больший размер и более гипокарниворный морфотип, а именно - большие и расширенные коренные и относительно короткий верхний хищнический зуб.
Позднеплейстоценовая форма из Палестины, N. vinetorum, также, возможно, была связана с N. megamastoiodes. Европейская форма вымерла в плейстоцене, в то время как N. sinensis эволюционировала в современный вид (Kurten, 1968).
Фоссилии из плиоцен-плейстоценовых отложений Южной Африки были описаны как N. terblanchei (Ficcarelli et al., 1984), N. lockwoodi из Эфиопии (Geraads et al., 2010) и N. abdeslami из Марокко (Geerards, 1997). Однако они отличаются дентальными характеристиками как от европейских форм, так и хронологически – между собой. Возможно, это свидетельствует о локальной эволюции.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#960 30 December 2018 16:34:27
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Подтриба Canina Fischer de Waldheim, 1817
Анализ группы Canini привёл к разделению американской клады Cerdocyonina, монофилетической группы, филогения которой была исследована Бертой (Berta, 1988) и Тедфордом и др. (Tedford et al., 1995). Остальные таксоны включают виды Canis и его сестринские таксоны, образующие терминальную кладу. Эта подтриба характеризуется следующими синапоморфиями: сильно изогнутая дорсовентрально скуловая арка и обычное наличие второго заднего бугорка на p4, расположенного между первым и цингулумом.
NUROCYON Sotnikova, 2006
РАСПРОСТРАНЕНИЕ: ранний плиоцен северо-восточной Монголии (Chonokhariakh-1)
ГОЛОТИП: полный череп (PIN 2737-257) с зубным рядом.
ДИАГНОЗ: отличается от Nyctereutes более крупным размером, более развитым лобным синусом, поперечно удлинёнными Ml-2 и наличием посторбитальных выступов без каких-либо признаков депрессии. Отличается от Eucyon наличием высокого черепа, крупного сагиттального гребня и пр. Отличается от плиоценовых видов Canis менее высоким черепом, более короткой лицевой частью, меньшим расширением затылочного гребня и др. Чрезвычайно удлинённая височная область и булла с почти плоской вентральной поверхностью являются дополнительными символами, отличающими Nurocyon от других Canini.
СОСТАВ РОДА: Nurocyon chonokhariensis Sotnikova, 2006
Nurocyon chonokhariensis (PIN 2737-257), голотип
ОБСУЖДЕНИЕ: демонстрирует необычную комбинацию примитивных и продвинутых черепных и дентальных особенностей. Хотя N. chonokhariensis значительно крупнее, чем ископаемые экземпляры Nyctereutes и достигает размеров и пропорций самых ранних представителей Canis, он близок к енотовидным собакам в большинстве дентальных признаков. В то же время сходство с Nyctereutes, по-видимому, можно интерпретировать как независимую тенденцию в развитии гиперкарниворных адаптаций. Почти все прогрессивные символы N. chonokhariensis являются синапоморфиями с группой Canis. В пределах этой группы монгольская собака достигает размеров современных шакалов и койотов, но пропорционально ближе к ископаемым формам вследствие более вытянутой и узкой посторбитальной области черепа.
Таким образом, N. chonokhariensis эволюционно более развит, чем самые производные формы Eucyon, однако имеет большее число плезиоморфных признаков, чем самые примитивные представители Canis. Филогенетически он, скорее всего, относится к отдельной линии с развитыми гиперкарниворными адаптациями.
XENOCYON Kretzoi, 1938
РАСПРОСТРАНЕНИЕ: позднего плиоцена и среднего плейстоцена Голарктики
ДИАГНОЗ: размер крупный, сравним с C. lupus; нижняя челюсть массивная, клыки укорочены, M1 со слабо развитыми цингулумами, M2 заметно не уменьшен по сравнению с M1; p4 с задним бугорком; m1 значительно редуцирован, m3 наличествует.
СОСТАВ РОДА:
СЕВЕРНАЯ АМЕРИКА:
Xenocyon texanus Troxell, 1915
РАСПРОСТРАНЕНИЕ: средний плейстоцен Аляски и Канады
ДИАГНОЗ: отличается от X. lycaonoides, m1 более широкий относительно в тригониде; гипоконид отделён от талонида и метаконида, М1 крупнее, чем у X. lycaonoides.
ОБСУЖДЕНИЕ: Берта (Berta, 1988) выделила texanus из рода Protocyon, ссылаясь на наличие метаконида на m2, несмотря на его исчезновение на m1. Она назначила этот таксон в род Cuon на основании гнатокраниальных отличий, однако она не заметила, что Х. texanus сохраняет m3 - характеристика, отсутствующая у Cuon.
Xenocyon (?) lycaonoides Kretzoi, 1938
РАСПРОСТРАНЕНИЕ: средний плейстоцен Аляски
ДИАГНОЗ: отличается от X. dubius более крупным размером; премоляры не столь удлинённые, а также более широкие и высокие; на p2 отсутствует задний бугорок; m2 с метаконидом, меньшим по сравнению с протоконидом и более узким талонидом.
ОБСУЖДЕНИЕ: размерно находится в нижнем диапазоне X. lycaonoides и морфологически близок с этой формой.
АЗИЯ:
Xenocyon antoni Zdansky, 1924
РАСПРОСТРАНЕНИЕ: поздний плиоцен - ранний плейстоцен Азии (Китай)
ОБСУЖДЕНИЕ: морфологически близок с европейским Х. falconeri[/i].
Xenocyon dubius Teilhard De Chardin, 1940
РАСПРОСТРАНЕНИЕ: средний плейстоцен Китая
ДИАГНОЗ: имеет специализацию нижнего хищнического зуба, соответствующую специализации m1 у X. lycaonoides. При этом у китайской формы имеется нередуцированный m2 и канидоподобный m3.
ОБСУЖДЕНИЕ: основные характеристики китайской формы позволяют отнести её к роду Хепосуоп. Различия в размерах у X. lycaonoides и X. dubius колеблются в пределах изменчивости одной популяции современного C. lupus и поэтому, по-видимому, целесообразно считать их одним видом.
ЕВРОПА:
Xenocyon falconeri Forsyth-Major, 1877
Xenocyon falconery ( IGF 865) Верхнее Вальдарно, Италия
РАСПРОСТРАНЕНИЕ: ранний плейстоцен Италии и Испании
ДИАГНОЗ: размер, сопоставимый с размерами крупнейших современных волков. Отличается от Canis ex gr. etruscus и Canis ex gr. arnensis как на основе крупного размера, так и брахиодонтности верхних моляров и хищнических зубов.
ОБСУЖДЕНИЕ: Понс Мойя (Pons Moya, 1987) предполагал синонимию X. lycaonoides с X. falconeri, но это решение слишком упрощено, так как изучение виллафранкских псовых позволяет отличать этот вид и более прогрессивные среднеплейстоценовые формы (Rook, 1993). Торре (Torre, 1967) также описывал специфические характеристики Х. falconeri, отмечая близость с Х. antonii.
Xenocyon lycaonoides Kretzoi, 1938
Xenocyon lycaonoides (VM-2255), Вента Мицена, Испания
РАСПРОСТРАНЕНИЕ: ранний плейстоцен Евразии
ДИАГНОЗ: размеры крупные. М3 всегда присутствует. m1 не редуцированы. Талонид на m1 куоноподобный с крупным гипоконидом. Метаконид на m1 небольшой, плотно прилегает к протокониду. Р4 широкий, массивный, со слабовыракенным протоконовым бугорком и удлинённой лопастью метакона. На М1 отчётливый внутренний бугорок за метаконом не сформирован.
ОБСУЖДЕНИЕ: по-видимому, эволюционировал из X. falconeri. Европейские образцы демонстрируют морфологию нижнего карнассиала, который имеет тенденцию к специализации.
АФРИКА:
Xenocyon africanus Pohle, 1928
РАСПРОСТРАНЕНИЕ: поздний плиоцен - ранний плейстоцена Африки
ОБСУЖДЕНИЕ: установлен на материале из Олдувэя как «Canis» africanus, в то время как Брум (Broom, 1948) описывал «Canis» atrox на материале из Кромдраая. В настоящее время эти виды считаются конспецифичными или, по крайней мере, тесно связанными (согласно Masini & Torre, 1990 и Torre et al., 1992). Не исключено, что к этой форме относится и «Canis» africanus из Южной Африки (Olduvai, Kromdraai). В то же время Тёрнер (Turner 1986, 1990) считал экземпляры из Kromdraai как Canis sp. без какой-либо связи с Xenocyon.
ВЫВОДЫ: крупные Canidae из позднеэоплейстоцен-раннеплейстоценовых отложений Европы частично относились к Canis, а также частично рассматривались как формы, близкие Lycaon. Кретцой (Kretzoi, 1938) дал этой форме новое родовое название - Xenocyon (описав нижнюю челюсть как X. lycaonoides, а верхние зубы - как C. gigas). Впоследствии Музиль (Musil, 1972) объединял остатки крупных Canidae из целой серии среднеевропейских местонахождений в один вид X. spelaeoides.
В плиоценовых фаунах Голарктики пока не найдено остатков канид, которые могли бы рассматриваться в качестве предковой формы для этого рода. Появление рода отмечается только с эоплейстоцена. О последующем развитии Хепосуоп мнения исследователей весьма противоречивы.
Образцы из позднего плиоцена Азии, а также те из плио-плейстоцена Европы характеризуются более широким талонидом на M1 и более широким М2 и рядом топологических отличий. Эта примитивная форма фиксируется в Китае как Х. antonii, а также в Европе - X. falconeri. Образцы из раннего плейстоцена - начала среднего плейстоцена Евразии и Африки характеризуются уменьшением талонида на M1, увеличением гипоконида, а также широким M2. Эта форма фиксируется как X. lycaonoides, X. africanus и пр.
Имеется гипотеза Торре (Torre, 1967), предполагающего, что существует супраспецифическая группа Xenocyon ex gr. falconeri, состоящая из таких видов, как Х. falconeri (ранний плейстоцен Европы) Х. antonii (поздний плиоцен и ранний плейстоцен Азии) и Х. africanus (поздний плиоцен и ранний плейстоцен Африки). Диагноз этой группы отличается от Xenocyon sensu stricto примитивным состоянием большинства дентальных признаков.
Также интересна группа псовых плио-плейстоцена о. Ява, описываемых как Cuon trinilensis, C. sangiranensis, Mececyon trinilensis и Megacyon merriami. Исследования показывают, что в раннем и начале среднего плейстоцена в этом регионе Cuon отсутствовал, поэтому Xenocyon – единственный претендент. Очевидно, что мигрировавшая в ци-саате с материка крупноразмерная форма (Megacyon) в триниле (начале среднего плейстоцена) сменилась небольшой (Mececyon). Последняя, вероятно, является следствием анагенетической островной карликовости (40% от начального размера) в результате конкуренции в гильдии хищников (van der Geer, 2018). В то же время остатки этих форм очень незначительны и не позволяют сделать какие-либо радикальные выводы относительно их происхождения, эволюции и филогенетических связей (Schtitt, 1973).
Тениус (Thenius,1954) считал Х. lycaonoides предком раннеплейстоценового Cuon priscus, но поскольку в настоящее время установлена одновозрастность находок обоих видов, вопрос об их филогенетической близости сомнителен.
Вероятная связь с современным родом [i]Lycaon, по-видимому, подтверждается наблюдениями по морфологии, в то же время Lycaon в отличие от Cuon и Canis характеризуется особенно длинным талонидом на m1, тогда как талонид Хепосуоп укорочен, как у куонов. Если бы Lycaon произошел от [i]Хепосуоп, то у него должно было бы наблюдаться вторичное удлинение талонида. Однако у Canidae преобладает тенденция к редукции талонида на m1, поэтому едва ли можно связывать происхождение Lycaon с Хепосуоп. Скорее всего, сходство зубных характеристик у этих двух родов - результат конвергентного развития, тем более что куоновая и ликаоновая специализация неоднократно проявлялась в различных филогенетических ветвях Canidae, например у кустарниковой собаки.
CYNOTHERIUM (сардинийский волк) Studiati, 1857
РАСПРОСТРАНЕНИЕ: эндемик среднего – позднего плейстоцена средиземноморских островов
ДИАГНОЗ: Нижние челюсти довольно массивные. Имелись диастемы p2/p3 и p3/p4, p2 бугорков. p3 и p4 с умеренно развитыми дополнительными вершинками, m2 с одновершинным тригонидом. C. malatestai отличался от С. sardous более крупным размером.
СОСТАВ РОДА:
Cynotherium malatestai (Madurell-Malapeira et al., 2015) – начало среднего плейстоцена Сардинии ( Италия )
Cynotherium sardous (Studiati, 1857) - средний – поздний плейстоцен Сардинии (Италия) и Корсики ( Франция ).
Cynotherium sardous (CB 84-8022)
ОБСУЖДЕНИЕ: в качестве возможного предка Cynotherium было предложено несколько материковых видов, что фактически является филогенетически возможным для любой евразийской собаки. Это по двум причинам: во-первых, сильное воздействие эндемизма скрывает филогенетические признаки, разделяемые с материковыми видами; во-вторых, время прибытия на остров столь же широкое, как и вся история канид в Евразии, поскольку Сардиния-Корсика неоднократно колонизировались с позднего миоцена до позднего плейстоцена. В связи с этим Ван дер Мэйдом (Van der Made, 1999) предложено несколько гипотез происхождения Cynotherium:
1. Линия Nyctereutes - соотношения и общий вид черепа Cynotherium напоминают пропорции Nyctereutes. Однако это сходство только поверхностно, так как имеются несколько краниальных особенностей - у Nyctereutes имеются сильные сагиттальные и парасагиттальные гребни, а также дробящая секция зубов у Nyctereutesгораздо более развита, чем режущая - ситуация, противоположная наблюдаемой у Cynotherium. Также у Nyctereutes отсутствует второй выступ на p3.
2. Линия Eucyon – возможность того, что Cynotherium является независимой эволюционной линией, происходящей от эуциноподобного предка, развившего канисоподобные характеристики в условиях длительной островной изоляции. Действительно, имеются черты, напоминающие Eucyon: небольшая лобная область, скуловые дуги, передние конечности относительно короткие и др. Однако, хотя эти признаки наблюдаются у примитивных представителей Canini, скорее всего, они являются результатом островной адаптации и, следовательно, представляют параллельное развитие.
3. Линия Canis (C. arnensis или C. mosbachensis) – эти представители имеют широкие морды, сильные и широкие скуловые арки и хорошо развитые сагиттальные гребни. У Cynotherium не хватает всех этих признаков: у него узкая морда, слабые и узкие скуловые дуги, очень низкий сагиттальный гребень и тонкая челюсть.
Эту загадочную комбинацию гиперкариноподобного зуба и гипокариновидного черепа можно объяснить следующим: Cynotherium происходит от гипо- или мезокарниворного предка и развивает гиперкарниворный зубной ряд независимо. Проблема здесь заключается в том, что никогда в истории канид у таксона не появляется гиперкарниворный дентальный морфотип без связанных с ним краниальных признаков. Следовательно, Cynotherium эволюционировал от предка с гиперкарниворным морфотипом, а гипокарниворные черепные характеристики приобрелись изоляционно. В этом случае морфология зубов Cynotherium отражает сохранение предковых характеристик. Известно три русцинийских группы: Xenocyon, Cuon и Lycaon. Поскольку современные и ископаемые Cuon лишены m3, а у Lycaon имеются развитые передние вершинки на p3-p4, отсутствующие у Xenocyon и Cynotherium, наиболее вероятным предком является Xenocyon.
Cynotherium malatestai (MPUR/V4200a), голотип
Это означает, что Xenocyon, должно быть, прибыл на Сардинию/Корсику где-то в конце раннего – начале среднего плейстоцена, потому что именно с этого времени X. lycaonoides регистрируется в нескольких евразийских местонахождениях.
CUON (красный волк) Hodgson, 1838
РАСПРОСТРАНЕНИЕ: средний плейстоцен (Евразия, Северная Америка) – ныне (Евразия)
ДИАГНОЗ: остеологическая дифференциация Cuon от Xenocyon и морфологически связанного Lycaon заключается главным образом в утрате m3; редукции m2, исчезновении энтоконида на m1 и пр. Принимая во внимание отличия, наблюдаемые в талонидном бассейне и, в частности, в гипокониде, можно констатировать, что Cuon явно отличается от Lycaon и Xenocyon, причём последний, возможно, является его предком (Tedford et al., 2009).
ВЫМЕРШИЕ ФОРМЫ:
Cuon javanicus antiquus (Matthew and Granger, 1923) - средний плейстоцен Азии
Cuon alpinus priscus (Thenius, 1954) – средний плейстоцен Европы
Cuon alpinus fossilis (Nehring, 1890) – средний-поздний плейстоцен Азии
Cuon alpinus caucasicus (Baryshnikov, 1978) – поздний плейстоцен Евразии
Cuon alpinus europaeus (Bourguignat, 1868) – поздний плейстоцен Европы
Cuon primaevus ssp. (Hodgson, 1837) – поздний плейстоцен Северной Америки
ОБСУЖДЕНИЕ: самые ранние формы Cuon известны из среднего плейстоцена Европы (C. alpinus priscus) и Юго-Восточной Азии (C. javanicus antiquus) Позднеплейстоценовые формы северной Азии обычно именуют как C. alpinus fossilis, европейские - C. alpinus europaeus. В ход описания фоссильного материала Адам (Adam, 1959) выводил эволюционную линию C. alpinus, начиная с C. a. priscus через C. a. fossilis - C. a. europacus, с прогрессивными модификациями моляров – потери метаконида на m2 и увеличении переднего бугорка на p4.
Ископаемая летопись Cuon в европейском регионе очень ограничена по сравнению с Canis mosbachensis или C. lupus. И, по-видимому, перекрытие трофических ниш было высоким. Из Северной Америки известны образцы из ранчолабрия северо-восточной Мексики – эта находка является напоминанием о скудости нынешних реальных представлений даже о позднеплейстоценовых таксонах. Не исключено, что популяции из северо-восточной Азии во время снижения уровня моря вполне могли пересечь Берингию и спуститься в более низкие широты, избегая конкуренции с Canis и Xenocyon.
LYCAON (гиеновидная собака) Brookes et al., 1827
РАСПРОСТРАНЕНИЕ: поздний плиоцен – ныне, Африка
ДИАГНОЗ: L. sekowei размерно средний между L. pictus и C. lupus; зубы относительно высококоронкованы, премоляры укорочены; р4 с крупным, но сниженным протоконом, m1 удлинённый, с широким бассейном. L. magnus чуть крупнее, отличается от типовой формы отсутствием передних вершинок на нижних премолярах и размером метакона на m1; нижние премоляры L. pictus меньше, чем у L. magnus,
Lycaon sekowei Hartstone-Rose et al., 2010
Lycaon sekowei (CD 8280/8281/8285), голотип
РАСПРОСТРАНЕНИЕ: поздний плиоцен (1,9 млн) Южной Африки
ОБСУЖДЕНИЕ: зубы этой формы более высокие, чем у Xenocyon или C. lupus. По-видимому, имел более широкое нёбо, чем у Xenocyon (как у L. pictus). Аналогично уменьшен буккальный цингулум на коренных зубах; а также, подобно Xenocyon и L. pictus, редуцированы m1 и m2.
Lycaon magnus Ewer and Singer, 1956
РАСПРОСТРАНЕНИЕ: средний плейстоцен (0,8 млн) Африки (Tighenif, Elandsfontein)
ОБСУЖДЕНИЕ: сходен с типовой формой редуцированным гипоконом на M1. Отличается от современного L. pictus крупным размером, очень уменьшенным метаконидом m1 и отсутствующим гипоконулидом. Образцы из Elandsfontein отличаются от L. pictus морфологией премоляров, но сходны в этом с экземплярами из Tighenif. Следовательно, в раннем и среднем плейстоцене L. magnus имел панафриканское распространение.
Lycaon pictus Temminck, 1820
Lycaon pictus ( RMCA 46B), Конго
РАСПРОСТРАНЕНИЕ: средний плейстоцен – ныне (Африка)
ОБСУЖДЕНИЕ: существуют некоторые дебаты по географическому диапазону Lycaon. Куртен (Kurten, 1968) предполагал, что его фрагменты найдены из позднеплейстоценовых местонахождений Европе, но Тениус (Thenius, 1972) и Малькольм (Malcolm, 1979) считают, что они принадлежат Canis. Если это так, то Lycaon всегда был ограничен африканским континентом. Также имеется челюсть из пещеры Хайоним (Израиль), демонстрирующая плезиоморфность в небольшом энтокониде на m1 – он крупный у X. falconeri, но полностью отсутствует у современном Lycaon. Плезиоморфность также подтверждается для m2 – все эти черты несут сильное влияние ксеноционовых псовых. Присутствие ликаона в Западной Азии в конце среднего плейстоцена вновь открывает вопрос о том, была насколько велика область, в которой эволюционировал Lycaon.
В Африке идентификация Lycaon сложна в связи с отличиями от раннеплейстоценового волка, C. africanus. Следует отметить, что «Canis» africanus в настоящее время синонимизируется с Xenocyon, что выглядит вполне разумным на основе имеющегося материала.
ВЫВОДЫ: Lycaon, Canis и Cuon являются близкими родственниками, имеющего общего предка, но эволюционное происхождение Lycaon до сих пор плохо изучено. В качестве альтернативы было высказано предположение о том, что предком для Lycaon могли являться некие куоноподобные собаки, основываясь на гиперкарниворных адаптациях обеих форм, однако это сходство может быть альтернативно объяснено коэволюцией и евразийское происхождение с последующей радиацией в Африку примерно 2 млн лет назад выглядит сомнительным.
В настоящее время лучшая гипотеза - это эволюция от позднеплиоценового X. falconeri или раннеплейстоценового X. lycaonoides, хотя следует отметить, что демаркация между этими двумя видами Xenocyon в настоящее время не вполне ясна (Martınez-Navarro and Rook, 2003). Впрочем, некоторые авторы утверждают, что X. falconeri Европы, X. antonii Азии и африканский. X. ajricanus следует включить в род Lycaon на основании морфологического сходства. Однако существуют некоторые различия в общем размере и в зубах – к примеру, у Lycaon отсутствует энтоконид в талониде m1.
В связи с описанием L. sekowei эволюция гиперкарниворного морфотипа несколько прояснилась: на первом этапе появились дополнительные вершинки (развитые в большей степени, чем у Xenocyon; на втором увеличились и заострились бугорки защечных зубов; на третьем гипертрофировались посткраниальные адаптации к курсориальной локомоции.
Два фактора могут свидетельствовать об эндемичности Lycaon как африканского рода. Во-первых, это численно редкая форма с низкими плотностями популяций в экосистемах, что значительно ограничивает его палеонтологическую фиксацию. Во-вторых, Canis lupus и Cuon alpinus, две других социальных хищника, занимали обширные районы северного полушария и ограничивали расширения популяций Lycaon на протяжении большей части плейстоцена и голоцена.
PROTOCYON Giebel, 1855
РАСПРОСТРАНЕНИЕ: средний - поздний плейстоцен Южной Америки
ДИАГНОЗ: Protocyon scagliarum отличается от P. orcesi и P. troglodytes с наличием р4 с передним бугорком и диастемой нижних премоляров. Также отличается от P. troglodytes наличием переднего бугорка на р4, соединённого с более редуцированным протоконом. P. orcesi отличается от P. troglodytes отсутствием m3 и m2 с поперечно узким талонидом.
СОСТАВ РОДА:
Protocyon troglodytes (Lund, 1838) - Бразилия, Боливия, Эквадор и северная Аргентина
Protocyon scagliarum (Kraglievich) - Аргентина
Protocyon orcesi (Hoffstetter 1952) - Аргентина
Protocyon tarijensis (Ameghino 1902) - Боливия
Protocyon troglodytes (MCPU-PV 148)
ОБСУЖДЕНИЕ: размером и дентальной морфологией близок с Theriodictis, но протокон на р4 более редуцирован и позиционируется более мезиально; гипокон на m1 более уменьшен, а внутренняя половина. Кроме того, m1 у P. scagliarum и P. orcesi демонстрируют более развитый и мезиодистально спозиционированный гипокон
LUPULELLA (чепрачный шакал) Hilzheimer, 1906
Эндемичный род южноафриканских псовых с современными видами L. adustus и L. mesomelas.
ДИАГНОЗ: размерно отличается от Vulpes и Nyctereutes, будучи в габаритном диапазоне шакалов. Данный таксон действительно очень сходен с шакалами, однако отличается более коротким сагиттальным гребнем и укороченный кристид между гипоконидом и энтоконидом на m1. Отличается от C. aureus менее треугольной затылочной областью, P4 короче относительно M1 и более удлинённым M1, чем у большинства представителей этого вида, а также рядом других признаков. Также отличается в пропорциях щечных зубов: P4 короче относительно M1, а p4 и m2 более длинны относительно m1.
Lupulella paralius Geraads, 2011
РАСПРОСТРАНЕНИЕ: прим. 2,5 млн. (поздний плиоцен) Марокко (Ahl al Oughlam)
ГОЛОТИП: AaO-4057, деформированный череп с щечными зубами
ДИАГНОЗ: среднеразмерная форма, сагиттальный гребень укорочен, затылочная поверхность колоколообразная, паракон на P4 выше метакона, p4 относительно m1 удлинён, m2 с узким талонидом и пр.
Lupulella mohibi Geraads, 2011
Lupulella mohibi (GDR-7966), голотип
РАСПРОСТРАНЕНИЕ: прим. 0,5 млн. (средний плейстоцен) Северной Африки
ГОЛОТИП: OH1-GDR – относительно полный череп с разрушенной базикраниальной областью
ДИАГНОЗ: среднеразмерная канида, сагиттальный гребень очень короткий. Третьи резцы маленькие, M1 скорее трапециевидный чем треугольный. Отличается от Nyctereutes более узкой затылочной областью и морфологией нижней челюсти, от C. aureus и L. mesomelas укороченностью секториальной части зубного ряда, от L. adustus узкой посторбитальной областью, M1 короче по отношению к его ширине, а p4 короче относительно m1 и m2.
ОБСУЖДЕНИЕ: ископаемые формы описывались как L. mesomelas latirostris и L. m. pappos. Диапазон размеров широкий, но не более, чем у современного C. aureus и меньше в пределах одного местонахождения, предполагая, что наличествовали региональные или темпоральные варианты. Укороченными m2 с довольно узким талонидом и длинными Р4 ископаемые формы больше походили на современные L. mesomelas и отличались от современного L. adustus небольшим гипоконом на M1, более длинном P4 относительно M1 и удлинённом p4 относительно m1.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#961 30 December 2018 16:50:55
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
CANIS Linnaeus, 1758
ДИАГНОЗ: синапоморфии, объединяющие Canis с Cuon, Lycaon и Xenocyon, при морфологическом анализе одновременно выглядят отличительными признаками: увеличение лобного региона; на p4 есть дополнительная вторая постериорная вершинка; на m1 энтоконид соединён с гипоконидом и пр.
В принципе, род Canis по отношению к вышеуказанным таксонам выглядит несколько примитивнее вследствие относительно крупных клыков и отсутствия таких гиперкарниворных дентальных признаков, как удлинённость метаконида или редукция энтоконида на m1 – m2; редукции гипокона на M1 – M2 и пр.
СЕВЕРНАЯ АМЕРИКА:
Canis edwardii Gazin, 1942
Canis edwardii (FAM 63100), ирвингтоний Аризоны
ГОЛОТИП: NMNH 12862, фрагментарный череп с неполным зубным рядом из среднего плейстоцена Аризоны
РАСПРОСТРАНЕНИЕ: бланко – ирвингтоний (ранний - средний плейстоцен) Северной Америки (Калифорния, Флорида, Аризона, Небраска, Техас).
ДИАГНОЗ: отличается от C. edwardii и C. ferox комплексом синапоморфических признаков, разделяющих их с койотами и волками – топологией вершинок на хищнических зубах. Также крупнее койотов и отличается от них как позицией сагиттального гребня, так и пропорцией в дистали передних конечностей.
ОБСУЖДЕНИЕ: эндемичная форма Северной Америки – в гипотезе Сяомина и Тедфорда является потомком койотоподобного Eucyon davisi к C. lepophagus до койота (возможно, даже линейная эволюция). Возможно, C. edwardii, C. latrans и C. aureus даже образуют вместе некую кладу.
Canis cedazoensis Mooser and Dalquest, 1975
ГОЛОТИП: ТММ 41536-41, частичная правая верхняя челюсть с P3 – M1 и M2
РАСПРОСТРАНЕНИЕ: ранчолабрий (поздний плейстоцен) Мексики
ДИАГНОЗ: отличается от C. aureus и C. thooides в морфологии M1 (относительно длины Р4 со сниженным гипоконом и внутренним цингулумом); альвеола М2 указывают на меньший размер относительно М1.
ОБСУЖДЕНИЕ: морфология предполагает экстерьер койотоподобной собаки с большими гиперкарниворными тенденциями, чем у C. thooides и C. feneus.
Canis feneus Tedford et al., 2009
ГОЛОТИП: (FAM 25515) нижнечелюстные ветви из позднего ирвингтония Небраски
РАСПРОСТРАНЕНИЕ: поздний ирвингтоний (конец среднего плейстоцена) Небраски
ДИАГНОЗ: отличается от C. thooides и C. aureus утоньшёнными нижними премолярами; р4 меньше по отношению к m1 (особенно в области метакона), m 1 с редуцированным парастилем, р4 с чётко выраженным дополнитеьным задним бугорком, m 2 с увеличенным параконидом и пр.
ОБСУЖДЕНИЕ: почти современник C. cedazoensis, демонстрирующий более мезокарниворные (скорее даже гипокарниворные) признаки или, может быть, даже гипокарнитивные признаки. Известен только по фрагментарным остаткам отдельных особей и, если диагноз поставлен правильно, его присутствие свидетельствует о том, что до сих пор нераскрытое разнообразие шакало/койотоподобных псовых оккупировало весь североамериканский материк ещё в раннем – среднем плейстоцене.
Canis thooides Tedford et al., 2009
ГОЛОТИП: FAM 63101, нижние челюсти с зубным рядом и посткраниаль из бланко Аризоны
РАСПРОСТРАНЕНИЕ: ранний плейстоцен (бланко) Аризоны
ДИАГНОЗ: отличается от C. lepophagus и C. ferox, будучи более сходным с прогрессивным Canis в широких премолярах (морфология р4), а также топологии коренных зубов. 
Canis thooides (AMNHM, 81041)
ОБСУЖДЕНИЕ: интересно рассмотреть койота/шакала как продукт раннеплейстоценовой дивергенции (бланко) – радиацию этих форм Canis в Северной Америке. Видимо, это целая эволюционная клада, начавшаяся с C. cedazoensis и C. feneus. Возможно, близкая к C. aureus и, видимо, столь же космополитичная, как и волк/собака – у Арамбурга (Arambourg, 1979) упоминается C. anthus primaevus = C. aureus из раннего плейстоцена Марокко.
Canis armbrusteri Gidley, 1913
Canis armbrusteri (NMNH 11886)
ГОЛОТИП: NMNH 7662, часть левой нижней челюсти с щечными зубами из позднего ирвингтония Мэриленда.
РАСПРОСТРАНЕНИЕ: ранний ирвингтоний – ранчолабрий (1,9 млн – 240 тлн); ранний – поздний плейстоцен
ДИАГНОЗ: отличается от C. lupus морфологией премоляров, от C. dirus – отсутствием некоторых гиперкарниворных признаков – к примеру, губного цингулума на М1.
ОБСУЖДЕНИЕ: Новак (Nowak, 1979) полагал, что C. armbrusteri эволюционировал от C. edwardii и эта дивергенция произошла в начале плейстоцена. C. edwardii действительно имеет сходные синапоморфии с волками и койотами - редуцированность метаконида на m2; иная топология премоляров и пр. Тем не менее, краниальные пропорции C. edwardii весьма отличны от C. armbrusteri, который, в свою очередь, близок с C. dirus и C. lupus.
Очевидно, C. armbrusteri представляет первую линию волкоподобных канин в северной Америке – гипотетически, несколько связанной с C. chihliensis.
Canis ferox Miller and Carranza-Castaneda, 1998
Canis ferox (FAM 49298), паратип
ГОЛОТИП: IGM 1130, почти полный, но сильно фрагментированный череп с зубным рядом и связанной посткраниалью из позднего хемфиллия Мексики.
РАСПРОСТРАНЕНИЕ: поздний хемфиллий – ранний бланко (прим. 5 млн – 0,2 тлн) Северной Америки
ДИАГНОЗ: расширенный лобный синус, увеличенный угловой отросток нижней челюсти, топология затылочной области – явная дифференция C. ferox от Eucyon и С. lepophagus.
ОБСУЖДЕНИЕ: по - видимому, C. ferox является прямым предком C. lepophagus – возможного предшественника современного койота C. latrans (Miller et al., 2015).
Из-за сильно раздробленного состояния этого черепа его сходство с монгольской собакой трудно установить, хотя их краниальный размер, по-видимому, совпадает полностью. Кроме того, они разделяют длинную и узкую область посторбитального сужения, небольшие альвеолы верхнего клыка и раздвоенный гипокон М1. С другой стороны, четкий треугольный контур надокиситального щита с острой и заостренной вершиной отличает C. ferox от N. chonokhariensis и позволяет идентифицировать его как истинного члена группы Canis.
[i] Canis lepophagus Johnston, 1938
Canis lepophagus (WTUC 881), голотип
ГОЛОТИП: WTUC 881, череп с зубным рядом из позднего плейстоцена Техаса
РАСПРОСТРАНЕНИЕ: ранний бланко (ранний плейстоцен) - Невада, Небраска, Техас, Канзас, Небраска, Калифорния, Нью-Мексико, Аризона, Айдахо, Флорида и северо-западная Мексика.
ДИАГНОЗ: прогрессивные признаки, отличающие C. lepophagus от E. davisi – больший размер и более массивный череп и челюсти; расширенная лобная пазуха; сагиттальный гребень выше; от C. latrans – более посторбитальное сужение и не столь явная гипертрофированная гиперкарниворность зубов.
ОБСУЖДЕНИЕ: в описании C. lepophagus Джонстон (Johnston, 1938) описывал этот таксон как более грацильным в общих пропорциях черепа и скелета, чем C. latrans. Хотя C. lepophagus сохраняет ряд общих черт с Eucyon, это более продвинутая канида, разделяющая немало прогрессивных признаков с более крупными видами Canis: расширение лобных пазух, дорсально суженная затылочная область, морфология нижней челюсти, зубов, посткраниали и пр.
Canis dirus Leidy, 1858
Canis dirus (FAM 25511M, FAM 25511, FAM 25509) ирвингтоний Небраски
РАСПРОСТРАНЕНИЕ:: поздний ирвингтоний – ранчолабрий Северной Америки (Калифорния, Небраска, Канада, Мексика, Венесуэла, Эквадор, Боливия и Перу)
ДИАГНОЗ: выделяется рядом аутапоморфий, связанных с гиперкарниворностью: исчезновение передненаружного цингулума на m2, значительное снижение метаконида и энтоконида на m1 – m2; редукция гипоконов на M1 – M2 и пр.
ОБСУЖДЕНИЕ: в 1974 году Новак, Куртен и Берта предположили, что C. dirus всё же не является близкородственным C. lupus, что позже подтвердили такие маститые специалисты по псовым, как Сяомин и Тедфорд, которые выдвинули гипотезу о происхождении C. dirus из C. armbrusteri. Ранний волк из Китая – C. chihliensis, возможно, был предком как C. armbrusteri, так и C. lupus. И неожиданное появление C. armbrusteri в раннем плейстоцене Северной Америки предполагает, что это был иммигрант из Азии.
Большинство окаменелостей с востока Северной Америки - C. d. dirus датируются от 125 000 до 75 000 тлн, но западные C. d. guildayi не только меньше размерно, но и более поздние темпорально - таким образом, возможно, что guildayi эволюционировал от dirus. К слову, в Южной Америке образцы C. dirus, относящиеся к позднему плейстоцену, были обнаружены вдоль северного и западного побережий, но не обнаружены в Аргентине - в районе, где найдены C. gezi и C. nehringi.
В 2009 году высказывалось предложение о том, что C. armbrusteri был общим предком как для палеарктических, так и для этих южноамериканских волков. Позднее даже выдвигались гипотезы, что C. dirus и C. nehringi были одним и тем же видом…
Анализ географических отличий показал разницу в кранио-дентальных особенностях подвидов «лютоволка» - C. d. guildayi и C. d. dirus – в пропорции посткраниали, как минимум: у «дируса» лапы были длиннее в диапазоне 10-15%.
Canis priscolatrans Cope, 1899
Canis priscolatrans (NMNH 7660), ранний ирвингтоний, Мэриленд
РАСПРОСТРАНЕНИЕ: поздний ирвингтоний - ранчолабрий (Калифорния, Колорадо, Небраска, Арканзас, Техас, Западная Вирджиния и Мэриленд)
ОБСУЖДЕНИЕ: Новак (Nowak, 1979) дал очень качественный и всесторонний обзор позднеплейстоценовых койотов, сравнивая ранних койотоподобные формы с современными. Согласно его исследованию, C. priscolatrans меньше современных форм и также демонстрирует высокую вариативность, но более робустный в общем экстерьере. Не исключено, что к этому кластеру относятся также C. irvingtonensis и «C.» texanus.
Вообще, C. priscolatrans напоминал C. rufus размерно и в пропорциях черепа, но с более сложной дентальной топологией.
Куртен полагал, что C. priscolatrans эволюционировал от C. lepophagus и представляет собой популяцию крупных койотов, которые были предками ранчолабрийских и современных C. latrans. Он отмечал, что C. arnensis Европы демонстрирует поразительное сходство с C. priscolatrans и они мог бы представлять голоарктическую популяцию шакалов/койотов. Новак с этим не согласился, считая, что C. priscolatrans является аналогом европейского C. etruscus. Позднее Куртен предположил, что C. priscolatrans и C. etruscus были частью группы, которая привела к C. lupus, но не был уверен в том, что они развились отдельно от C. lepophagus или возможного общего предка, который от него эволюционировал.
Canis lupus Linnaeus, 1758
ОБСУЖДЕНИЕ: морфологически среднеплейстоценовые находки очень схожи с современными и, если геологическая атрибуция этого материал верна, их возраст составляет ок. 1,0 млн.
ЮЖНАЯ АМЕРИКА:
Canis gezi Kraglievich, 1928
РАСПРОСТРАНЕНИЕ: энсенадий (конец раннего – средний плейстоцен) Бразилии
Canis nehringi Ameghino, 1902
РАСПРОСТРАНЕНИЕ: луханий (средний – начало позднего плейстоцена) Аргентины
ОБСУЖДЕНИЕ: являются сестринскими или производными таксонами, соответствующими позднему бланко и ирввингтонию Северной Америки. Учитывая их сходства и хронологию, было предложено, что C. gezi был предком как C. nehringi, так и позднеирвингтонского С. dirus nebrascensis (предполагаемый ранний «лютоволк»), хотя C. nehringi более схож с ранчалобрейским C. dirus. Возможно, C. armbrusteri являлся общей предковой формой для обеих клад.
ВЫВОДЫ: самые ранние остатки псовых Северной Америки, которых однозначно можно отнести к роду Canis, фиксируются в позднем хемфиллии. В то же время, возможно, C. ferox морфологически является промежуточным звеном между Eucyon davisi и C. lepophagus. Хотя C. ferox очень схож с европейским C. arnensis, в Азии эта группа, вероятно, появилась несколько раньше, чем в Европе (Rook & Azzaroli, 1996).
Джонстон (Johnston, 1938) изначально описывал и считал C. lepophagus предшественником современного койота. C. lepophagus отличается от C. latrans примитивными характеристиками - меньшим размером, более узкой мозговой областью, узкой областью посторбитального сужения и менее высокими буллами. В то же время C. lepophagus обладает некоторыми особенностями группы Canis, которые отсутствуют у азиатских форм - удлинённой лицевой частью черепа, почти трёхугольной формой надокиситального щита и относительно узким и длинным P4.
В среднем плейстоцене североамериканские собаки стали крупнее и C. priscolatrans, видимо, дивергировался от крупного C. armbrusteri. Мартин считал, что C. armbrusteri был связан с волком, в то время как Новак полагает, чтобы C. armbrusteri ближе к [i]C. priscolatrans, который затем породил C. dirus. Тедфорд предположил, что [i]C. armbrusteri является общим предком C. gezi, C. nehringi и C. dirus – на основе того, что морфологически китайский плиоценовый волк C. chihliensis, возможно, был предком как для C. armbrusteri, так и для C. lupus до их миграции в Северную Америку.
АФРИКА:
Диагностически, найденные на территории африканского материка (Алжир, Эфиопия, Кения, Марокко, Южная Африка, Танзания, Замбия) ископаемые образцы рода Canis в большей степени принадлежат шакалам и ликаону. Самая ранняя находка - Canis sp. из верховьев южной Турквел (Turkwel) ок. 3,5 млн лет. Современные африканские представители рода являются примитивными и очень адаптированными формами, что объясняет их относительный дефицит в палеонтологической летописи. Особый интерес представляет m2 из местонахождения Aın Boucherit, принадлежащий африканскому золотистому волку Canis anthus, описанный как С. а. primaevus (Arambourg, 1979), сходный с африканским L. paralius и европейскими C. accitanus, C. adoxus и C. aureus, однако некоторые отличия исключают явную идентификацию с этими видами.
ЕВРАЗИЯ:
Наличествуют несколько таксонов, требующих радикальной таксоно-систематической «привязки», если можно так выразиться.
Canis adoxus Martin, 1973 – в морфологии коренных зубов более сходных с родом Vulpes. В настоящее время очевидно, что C. adoxus не является представителем группы Canis. Тем не менее, определение C. adoxus в Eucyon также сомнительно - отсутствие поперечного гребня на талониде m1, увеличенный I3, хорошо расширенный лобный синус – всё это сходно с Nurocyon. Скорее всего, N. chonokhariensis и C. adoxus являются сестринскими таксонами и не исключено, что его следует отнести к новому роду.
Испанская форма Canis cipio Crusafont, 1950 – ну, здесь даже хронологически (туролий) с канисами никакой связи быть не может.
Эта форма отличается от Eucyon в пропорциях верхних зубов, структуры M1 и в большем размере. По сравнению с Nurocyon, C. сipio также демонстрирует поперечно расширенные верхние моляры размером, равным размеру монгольской формы. Верхние моляры Nurocyon меньше, чем у C. cipio, но разделяет с ней такие примитивные черты, как крупный протокон и выраженные внутренние цингулумы. С другой стороны, C. сipio очень схож с Canis морфологией верхних коренных зубов.
Canis arnensis del Campana, 1913
ГОЛОТИП: IGF 867, череп и нижняя челюсть от двух особей
МЕСТОНАХОЖДЕНИЕ: верхнее Вальдарно, Тоскана (Италия), ранний плейстоцен (1,5 млн).
ОБСУЖДЕНИЕ: этот таксон был описан на основе многочисленных образцов и, хотя увязывался как с шакалами (Тоrrе, 1967) или «койотоподобными собаками» (Kurten, 1974), формально так и не диагностировался.
Canis senezensis Martin 1973
МЕСТОНАХОЖДЕНИЕ: два верхнечелюстных фрагмента из Сенез (Франция), ранний плейстоцен (2,0 млн).
ОБСУЖДЕНИЕ: исследование, основанное на образцах C. arnensis, показало, что пропорции и морфология зубов C. senezensis подтверждают морфологический тип этого таксона как ранняя форма C. arnensis. Однако это оспаривается рядом исследователей и его таксономический статус остается спорным.
Canis accitanus Garrido and Arribas, 2008
Canis accitanus (FP1-2001-0434), голотип
ГОЛОТИП: FP1-2001-0434 – череп с зубными рядами
МЕСТОНАХОЖДЕНИЕ: Фонелас (Испания), ранний плейстоцен (1,9-1,7 млн)
ДИАГНОЗ: небольшая собака с малоразвитым сагиттальным гребнем. Общая архитектура черепа и дентальные характеристики отличают его от Eucyon или Vulpes. Тем не менее, морфологические характеристики черепа и зубов аналогичны представителям рода Canis.
ОБСУЖДЕНИЕ: сравнение черепа C. accitanus с C. lupus, C. latrans, C. adustus, C. mesomelas и C. simensis показывает, что он имеет явные видовые особенности.
Canis etruscus Major, 1877
Canis etruscus, Италия: A. - IGF 12867 (AMNH 89779), голотип; B–E. - IGF 12334 (AMNH 89777)
ГОЛОТИП: IGF 12867, череп
МЕСТОНАХОЖДЕНИЕ: верхнее Вальдарно, Тоскана (Италия), ранний плейстоцен (1,5 млн).
ДИАГНОЗ: описавший этот таксон Мэйор (Major (1877) не предоставил формального диагноза, но последующие исследователи, анализировавшие ископаемых псовых из Тосканы, указали на средний размер, удлинённую лицевую часть черепа и некоторые дентальные особенности премоляр-молярного ряда.
ОБСУЖДЕНИЕ: Торре (Torre, 1967) признал синонимию [i]C. majori/C. olivolanus с С. etruscus, однако как и в описании C. arnensis, выдал, в основном, лишь метрические определения, что изначально создавало некоторую путаницу. К примеру, образцы из Эскель (Франция), приписываемая Бонифэем (Bonifay, 1971) к C. etruscus, расценивается как промежуточная форма между C. etruscus и C. lupus у Торре, в то время как Сотникова (2001) относит их к мосбахским волкам. На самом деле, упрощение понятий о C. etruscus и C. arnensis как о волко- и шакалоподобных собаках соответственно, не всегда верно, потому что C. arnensis больше похож на C. lupus, чем C. etruscus в отношении некоторых краниальных признаков, в то время как C. etruscus, очевидно, проявляет более широкий набор вариативных особенностей.
Недавние генетические исследования подтверждают, что этрусский волк представляет собой ключевой таксон и является самым старым «истинным» представителем рода Canis в Европе.
Одновременно в Европе встречались и такие волки, сведённые с ним в синонимию. Они были описаны под разными названиями - С. olivolanus или гомбасцогский С. strandi. Вообще, этрусский волк широко распространён по всей Европе, начиная с раннего виллафранка и обычен в лесных фаунах (Оливоль, Вальдарно, Эрпфинген), но редок в степных. Здесь интересно отметить, что с ним сближают С. falconeri из позднего виллафранка Италии (Вальдарно) - с крупными зубами, позволяющими предполагать у него преимущественное питание падалью. Возможно, что в систематическом отношении он был близок к С. dirus (Kurten, 1968).
Canis apolloniensis Koufos and Kostopoulos 1997
МЕСТОНАХОЖДЕНИЕ: пещ. Петралона (Греция), ранний плейстоцен
ДИАГНОЗ: небольшого размера; диастема между премолярами отсутствует; выраженный и хорошо разделённый протокон; М1 длинный с относительно коротким тригонидом.
ОБСУЖДЕНИЕ: сравнительно недавнее исследование (Geraads, 2011) предположило наличие двух флогенетических линий: одна линия - C. arnensis, C. accitanus, C. senezensis, другая - C. etruscus, C. appoloniensis. Действительно, диагноз Коуфоса и Костопулоса (1997), слабо оправдывает возведение нового таксона. В то же время выделение этого вида согласуется с мнением Рука и Торре (Rook and Torre, 1996) о наличии терминальной виллафранкской формы Средиземноморья, отличающейся от типичного C. arnensis.
Canis mosbachensis Soergel, 1925
ГОЛОТИП: так и не выбран из коллекции в составе пяти нижнечелюстных образцов, изученных Зоргелем в 1925 году в музее Майнце.
МЕСТОНАХОЖДЕНИЕ: Мосбах (Германия), начало среднего плейстоцена
ДИАГНОЗ: мелкоразмерные собаки – менее современного C. rufus. Тениус и Арго считают C mosbachensis подвидом C. lupus, однако другие исследователи не видят ясной связи между C. mosbachensis и C. etruscus полагая, что C. mosbachensis более похож на C. arnensis и что он имеет размер и дентальную морфолгию, более схожую с шакалоподобными псовыми.
ОБСУЖДЕНИЕ: Зоргель в сравнительном исследовании выделял из этого таксона форму C. neschersensis, Тениус (1954) считал его примитивным C. lupus, Сотникова (2001) считает его полноценным таксоном видового ранга из среднего плейстоцена Германии.
Остатки мелкоразмерных волков, также определяемых как С. mosbachensis, древнейшие находки которого относятся к «до-кромер»-ской эпохе (700 тлн) из Германии (Мосбах, Мауэр, Зюссенборн, Фойгтштедт), Австрии (Хундсгейм) и Венгрии относятся к С. etruscus. Однако в этом случае этрусский волк доживает в Европе до минделя. Размеры зубов у обоих форм действительно перекрываются и найти достоверные различия между ними, учитывая большую вариабельность дентальных признаков затруднительно, но при более широкой трактовке С. etruscus мелкий С. mosbachensis представляет собой отдельную, менее специализированную эволюционную ветвь.
Canis variabilis Pei, 1934
ЛЕКТОТИП: большая часть оригинальных образцов утрачена во время 2-й Мировой, в результате чего именование variabilis стабилизируется на основе гиподигмы из коллекций 1934 года.
МЕСТОНАХОЖДЕНИЕ: Чжоукоудянь 1 (Zhoukoudian), средний плейстоцен Китая
ДИАГНОЗ: обладает более короткими носовыми костями и изменениями в морфологии верхних коренных зубов. Череп отличается от типичного волка гораздо меньшим размером с более грацильной лицевой областью и заметно уменьшенным сагиттальным гребнем. Кроме того, нижняя граница некоторых челюстей C. variabilis сильно выпуклая, как и у С. familiaris.
ОБСУЖДЕНИЕ: краниодентальные характеристики более прогрессивны и указывают на то, что он был более гиперкарниворен, чем C. chihliensis, C. etruscus, C. arnensis, но менее, чем C. lupus. Он не является прямым предком волка, но был близким родственником.
Нынешние исследования предполагают, что именно C. variabilis может быть предком современной собаки (Wang, 2012). Анализ Уонга показал, что C. variabilis был «очень странным» в сравнении с другими китайскими Canis, поскольку он имел гораздо меньшие черепно-зубные размеры, чем C. lupus. Гипотетическое сходство с C. familiaris несколько подтвердил и анализ митохондриальной ДНК в 2015 г., подтвердив немаловажный вклад в родословную современной домашней собаки от этих псовых.
Canis antonii Zdansky, 1924
Canis antonii (FAM 97052), Нихэвань (Китай)
ГОЛОТИП: M3514, фрагментарный череп с зубным рядом
МЕСТОНАХОЖДЕНИЕ: в раоне дер. Яншао округа Миньчи, провинции Хэнань (восточно-центральный Китай), ранний плейстоцен.
ДИАГНОЗ: отличается от большинства канид менее хорошо развитым протоконом на P4 и увеличенным M1. Хотя C. antonii и C. chihliensis сопоставимы по размеру, C. chihliensis имеет более массивный протокон на P4 и М2, который меньше снижен относительно такового на M1, подразумевая, что этот вид является более примитивной формой.
ОБСУЖДЕНИЕ: родственный таксон группы C. armbrusteri, отличаясь от таковой аспектами дентальной морфологии, сохраняющей плезиоморфные особенности в топологии премоляр-молярного ряда зубов верхней и нижней челюсти.
Canis chihliensis Zdansky, 1924
Canis chihliensis (IVPP V 18333), Нихэвань
ГОЛОТИП: M3496, правый верхнечелюстной фрагмент с премоляр-молярным рядом.
МЕСТОНАХОЖДЕНИЕ: провинция Чихли (ныне Хэбэй), ранний плейстоцен.
ДИАГНОЗ: крупноразмерная собака, сагиттальный гребень высокий и длинный, I3 массивный, P4 удлинённый и с хорошо развитым протоконом, M1 мезиодистально сжат, m1 с метаконидом и энтоконидом, M2 и m2 крупнее относительно M1 и m1 соответственно.
ОБСУЖДЕНИЕ: де Шарденом и Пивето (Teilhard de Chardin and Piveteau, 1930) признавалось три морфотипа ископаемых канисов из Нихэвань – C. chihliensis (типовая форма), C. chihliensis var. palmidens и C. chihliensis var. minor.
Однако эти обозначения были поставлены под сомнение рядом последующих авторов. Так C. chihliensis palmidens был повышен до уровня вида (Tedford et al., 2009), некоторые образцы C. chihliensis из Loc.18 обозначены как C. teilhardi (Qiu и др., 2004). Два зуба из Danangou – Р4 (IVPPV 7252-1) и M1 (V 7252-2), первоначально идентифицированные как C. c. minor слишком малы, чтобы быть включёнными в этот список. Кроме того, протокон на P4 слишком уменьшен и расположен слишком далеко впереди, а лингвальная область M1 очень узкая – скорее всего, эти зубы принадлежат представителю рода Vulpes.
Canis palmidens Teilhard de Chardin and Piveteau, 1930
ЛЕКТОТИП: TNP 00198, череп
МЕСТОНАХОЖДЕНИЕ: Нихэвань (ныне Хэбэй), ранний плейстоцен.
ДИАГНОЗ: значительно меньшего размера, чем C.chihliensis с пропорционально явно более крупными премолярами.
ОБСУЖДЕНИЕ: в то время, как его авторы указывали на близкое сходство с C. arnensis и C. nescherensis, предполагая, что таксономическое выделение C. palmidens основывалось главным образом на его географическом положении, Торре (Тоrrе, 1976) придерживался несколько другой точки зрения с учётом хронологических диапазонов. Согласно анализу Гераадса (Geraads, 2011), образцы C. palmidens из Нихэвань вполне достойны выделения в отдельный видовой кластер.
Canis teilhardi Qiu, Deng and Wang, 2004
ГОЛОТИП: HMV 1144, череп и ассоциированная левая плечевая кость
МЕСТОНАХОЖДЕНИЕ: восточный склон Лонгдань, 2,5 км к югу от Валеси, провинция Ганьсу (северный Китай), начало раннего плейстоцена
ДИАГНОЗ: очень сходен с C. chihliensis, однако отличается менее прогрессивным морфотипом Р4, M2 и m2 меньше по сравнению с размерами M1 и m1 соответственно, и выраженным метаконулем на M1.
ОБСУЖДЕНИЕ: имеется широкая выборка из типового местонахождения (остатки 40 особей), подтверждая таксономическую валидность этой формы. Морфологический анализ ставит C. teilhardi в качестве сестринского таксона к C. chihliensis.
Canis vitastensis Kotlia, 1987
Canis vitastensis (VPL/B 3001), голотип
ГОЛОТИП: VPL/B 3001, фрагмент левой нижнечелюстной ветви с m2.
МЕСТОНАХОЖДЕНИЕ: палеорусло в 12 км от и Кашмира, средний плейстоцен
ДИАГНОЗ: ветвь стройная, параконид на m2 слабо развит, чем отличается от C. curvipalatus и C. cautleyi; метаконид остроконечный; протоконид высокий; гипоконид развит плохо; энтоконид ниже гипоконида; талонид треугольный и небольшой; тригонид меньше талонида; m3 наличествует.
ОБСУЖДЕНИЕ: соотношение переднезаднего диаметра зубов сходно с Canis и отличается от представителей Amphicyoninae. Кроме того, m2 легко отличается от других видов Canis, имеющих более развитый передний цингулум (по сравнению с параконидом) и обладают округлым энтоконидом - более высоким, чем гипоконид.
Canis tamanensis Vereshchagin, 1957
ГОЛОТИП: PIN 1249/621, нижнечелюстной фрагмент с карнассиальным рядом
МЕСТОНАХОЖДЕНИЕ: Синяя Балка (Краснодарский край), начало раннего плейстоцена
ДИАГНОЗ: отличается от C. lupus меньшими размерами, более высоким и прямым положением p4 относительно m1 и наличием заднего базального цингулума в p4 - эти признаки объединют его группой таких канид, как C. etruscus, C. arnensis и C. mosbachensis. Однако имеются отличия в более сниженной позиции p4 относительно m1 и несколько отличающейся морфологии коренных зубов. В целом, C. tamanensis хорошо соответствует характеристикам C.mosbachensis.
ОБСУЖДЕНИЕ: при создании C. tamanensis Верещагин (1957) указал на сходство новой формы с современной группой куонов-ликаонов ввиду отсутствия m3. Современные исследования всё же указывают на некоторое сходство с мелким европейским волком C. mosbachensis. Принимая во внимание аберрантность структуры m1 у других ископаемых канид этого временного интервала и отсутствие других образцов, C. tamanensis остаётся валидным, пока не будет доступен дополнительный материал о псовых из этой области.
Canis aureus Linnaeus, 1758
Наиболее ранние находки происходят из среднего и позднего плейстоцена Северной Африки и Италии. Из позднего плейстоцена Палестины определены остатки своеобразного С. lupaster (Kurten, 1965). Этот обитатель аридных ландшафтов встречается в Египте и Передней Азии. В систематическом отношении его сближают то с С. aureus (Ellerman, Morrison-Scott, 1951), то с С. lupus (Ferguson, 1981) – хотя, скорее всего, он является самостоятельным видом. К тому же это подтверждает, что шакалы не составляют однородную генетическую группу, несмотря на их большое морфологическое сходство.
В Европе ископаемые остатки мелких псовых известны из Италии. Франции, Германии, Австрии, Венгрии и т. д., - их обычно сближали с шакалами (Freudenberg, 1914). Последующие исследования показали, что большинство их них относится к некрупным плейстоценовым волкам – к примеру, С. mosbachensis. Также как и С. arnensis также долгое время относили к группе шакалов.
На Азиатском материке шакало-(койото)подобным формам принадлежат ископаемые остатки из Таджикистана (Куруксай), Забайкалья (Береговая), Монголии (Шамар, Диске) и Китая. Таким образом, койоты, являющиеся в настоящее время эндемиками североамериканского континента, в плейстоцене встречались по всей северной половине Голарктики.
По большому счёту, отчётливо наличествуется значительная обособленность группы койотов/шакалов, подтверждая воззрения на их подродовой статус (Огнев, 1931), так как эволюционно явно выделяются две плейстоценовые линии развития волкоподобных и койото/шакалоподобных собак.
Canis lupus Linnaeus, 1758
Волки появляются в Европе примерно в среднем плейстоцене (миндель-рисское межледниковье) – фиксируется в Люнель-Вьель южной Франции.
Центр происхождения вида установить сложно ввиду высокой вариативности морфологических признаков, обилию современных подвидов, обширностью современного ареала и его исключительной эвритопностью. Тем не менее, при сопоставлении названных критериев при обзоре палеонтологических сведений есть основание считать, что родина волка находилась в пределах юга Восточной Европы и Передней Азии.
В морфологической эволюции волка подмечена тенденция в развитии прогрессивного хищнического типа строения черепа и зубной системы - происходило относительное укорочение ряда зубов за счёт утеснения предкоренных, сокращения длины мозговой капсулы и увеличения длины височного сужения (Верещагин. 1951). В то же время мнения современных палеонтологов о родословном древе волка разноречивы.
В современных исследованиях по истории четвертичных волков Евразии разными исследователями описано свыше десятка ископаемых форм, различия между которыми нередко не превышают диапазона индивидуальной или половой изменчивости потому сомнительно может быть поставлена в ряд эволюционных изменений рода Canis. Ещё Фрейденберг (Freidenberg, 1914), основываясь на европейских палеонтологических материалах, утверждал, что остатки мелких волков типа C. neschersensis, С. kronstadtensis свойственны раннему плейстоцену, а остатки крупных волков группы С. lupus появляются лишь в среднем. Русскими учёными установлено, что от грузинских (С. tengisii) бинагадинских (С. l. apscheronicus) и кавказских (С. l. cubanensis) волков ископаемые формы отличались меньшими размерами и рядом архаичных признаков (индексом талонида, формой метаконида на m1, его строением и т. д.).
Очевидно, что эти формы видовой, или чаще подвидовой, таксономической значимости являются большей частью результатом морфометрических упражнений на фрагментарном материале. Впрочем, это отнюдь не отрицает реального существования палеогеографической изменчивости популяций на протяжении плейстоцена. Очевидно, напросто широкие географические связи популяций обусловили нивелировку некоторых дивергенций, стимулированных зональными условиями обитания.
ВЫВОДЫ: в первую очередь при рассмотрении рода Canis следует узреть проблемы, возникающие при идентификации C. arnensis в качестве койота. Торре (Torre, 1967) провел исследование по верхним коренным зубам, показав, что шакалы и волки отличаются этими характеристиками и что в этом признаке C. arnensis сходен с шакалами. Это действительно так, но койоты и шакалы не сходятся в этом признаке (за исключением крупных образцов) и C. arnensis близки с койотом. Точнее, C. arnensis демонстрирует очень большую схожесть с ирвингтонийским C. priscolatrans, но азиатский материал, соединяющий две популяции ещё очень скуден. Будущие исследования могут подтвердить преподложение, что [i]C. arnensis и C. pristolatrans являются частями единой голарктической популяции койотов и в этом случае C. arnensis следует рассматривать как синоним или подвид C. priscolatran. Энигматичная азиатская форма, которая может оказаться связующим звеном в цепи популяции койотов – C. chihliensis. Тем не менее, доступные данные указывают, что эта форма может быть несколько более крупнее и, возможно, более тесно связанной с C. etruscus.
Средний плиоцен («волчий исход») – радиация представителей рода Canis в Азии происходила значительно раньше, чем в Европе. Появление примитивного С. sр. [i]etruscus фиксируется в Китае (Юше), примерно 3,4 млн лет. Материал представлен черепом FAM-97050, который отличается от C. etruscus более развитой лобной областью и более крупными замыщелковыми отростками, в то время как щечные зубы не превышают таковые у C. etruscus и C. mosbachensis. Первое появление истинных Canis в Европе близки к форме C. arnensis (3,1 млн) – следовательно, формы размером с волка появились на евразийском континенте практически одновременно в раннем виллафранке.
Поздний плиоцен - диверсификация Canis в позднем плиоцене Азии (2,6 млн) - самые распространённые и разнообразные коллекции известны из северо-западного Китая – C. teilhardi, C. longdanensis, C. brevicephalus. Таким образом, все базовые компоненты Canis плейстоценовой фауны Евразии уже «сложились» к позднему плиоцену на востоке Евразийского континента – койото- и волкоподобные формы, демонстрирующие гиперкарниворный морфотип.
Следующая значимая дата – центральная Азия и восточная Европа. Куруксай (Таджикистан) - небольшая форма [i]C. kuruksaensis, чья морфология несколько похож на C. brevicephalus из Лонгдана, отличаясь меньшими хищническими зубами и примитивной морфологией коренных зубов. Другой важный случай фиксации каниса размером с волка регистрируется в хапровской фауне южной России – из Хапров и Ливенцовки. Размер зубов хапровского волка находится в пределах верхнего диапазона C. arnensis, верхние зубы морфологически ближе всего к C. senezensis, в то время как широкий и короткий талонид на m1 больше схож с куруксайским, чем с C. arnensis или C. etruscus.
Волко/койото-размерные каниды плио-плейстоцена Западной Европы являлись представителями групп C. etruscus и C. arnensis и их родственными формами - C. accitanus и C. apolloniensis. C. etruscus датируется примерно ранним плейстоценом и отличается рядом плезиоморфных особенностей - массивностью лобной области, относительно короткой областью посторбитального сужения, позицией нижних премоляров относительно коренных зубов и т. д. И здесь следует обратить внимание на Кавказский регион - возможно, что кавказские волки этого периода представляют собой отдельную линию плио-плейстоценовых представителей рода Canis и именно эту территорию следует рассматривать в поисках родословной C. lupus. В плейстоцене группа азиатских волков продолжала включать в себя формы, обладающие как гипо-, так и гиперкарниворным морфотипом – C. antonii, C. chihliensis и др.
К концу раннего плейстоцена разнообразие видов волка заметно уменьшилось - преобладающими формами были C. mosbachensis и C. variabilis. Ассоциация этих хищников была широко распространена в Евразии от Англии до Греции в Европе и от высоких широт Сибири до Китая и Таджикистана в Азии.
Ну, пока что всё...
Конечно, наибольший «афган» с родом Canis. И хотя я не сторонник огульной таксономической консолидации, но здесь мне несколько непонятно – почему до сих пор хотя бы современные представители этого рода не централизованы в подроды типа волко-шакало-койотоморфных собак? Ведь тогда бы насколько Гордиев узел этой систематики оказался хоть частично развязан… Спасибо, хоть африканскую линию чепрачных шакалов в отдельный таксон вычленили. А по большому счёту, койото-шакалы всё же должны быть хоть подродом обозначены.
Ещё раз всех с 2019-м!!!
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#962 30 December 2018 22:26:08
- Miracinonyx
- Любитель животных

- Зарегистрирован: 05 December 2006
- Сообщений: 19226
Re: Ископаемые псовые (Canidae)
Потрясающая работа проделана, ты герой, Игорь!
Спасибо тебе огромное за информативнейший обзор семейства, читать не перечитать! )
И иллюстрации классные, поклон Роману!
Неактивен
#963 30 December 2018 23:39:11
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5945
Re: Ископаемые псовые (Canidae)
Ребята, на самом деле с поста с 948.
Вот теперь собачек более-менее знаю.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#964 04 April 2019 21:55:44
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
У кого-то имеются промеры черепов так называемых берингийских и пещерных волков? Берингийский волк представлял собой плейстоценовую экоморфу серого волка, размерами такой же, но с усилиенными челюстями (более широкими) и более крупными хищническими зубами.
С одной стороны такое ощущение, что серый волк в плейстоцене был забитым падальщиком. С другой стороны, добыча плейстоценовых серых волков в среднем была примерно такой же величины, что и у смилодонов - бизоны, крупные олени, овцебыки, лошади и т.п.
В Индии действительно волки не являются главенствующими хищниками и в качестве охотника на крупных эивотных дхоль там имеет все преимущества. Но на севере, по-моему, с момента своего появления всегда правили псовые центрального рода (по крайней мере родственные ему).
Хотелось бы еще сказать по поводу социальности этих групп псовых. Везде пишут, что дхоли создают более многочисленные стаи. Но по факту, серые волки могут создавать стаи по 20, а то и более особей. Все зависит от обилия пищи. Но, при одинаковых природных ресурсах дхоли должны создавать более многочисленные стаи, я думаю, так как сами они намного мельче (один и тот же участок моет прокормить одно крупное животное или два мелких).
Интересно все же, какую нишу занимал волк в плейстоцене, например Северной Америки. Начиная с голоцена, как ни крути, он является верховным хищником этого континента.
Отредактировано Crazy Zoologist (04 April 2019 22:06:10)
#965 12 April 2019 17:07:20
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Ископаемые серые волки. Верхний плейстоцен Южной Италии. Шкала- 5 см.


Сколько см в длину крупнейший из известных черепов волка? Вот достаточно крупный, более 26 см. Плейстоцен Флориды.
Как я уже писал, у плейстоценовых волков челюсти были немного шире, а хищнические зубы массивнее, чем у современных. Размеры как у крупных современных, то есть в среднем намного больше. Но, справедливости ради, надо сказать, что у гиеновой собаки и челюсти и хищнические зубы еще более усилены, по сравнению с волчьими. Если смотреть на краниодентальные характеристики, то ликаон получается более падалеяден (костедробилен), но на практике это не так. Волки в среднем убивают животных такой же весовой категории, а некоторые популяции валят и более крупных животных, чем гиеновые собаки. Зимой волки едят падали, я думаю, поболее, чем ликаоны и утилизируют более крупные кости. Трухлявого лося утилизируют в Евразии в первую очередь волки, а в Африке трупы животных такого размера достаются лишь более крупным хищникам, ну и, конечно же, птицам-падальщикам.
Волк убиваетболее крупных животных и разгрызает более крупные кости, нежели ликаон, лишь по причине более крупных размеров. Ликаон в Африке занимает нишу небольшого коллективного хищника и в более тяжелую весовую категорию не лезет. Кстати, волки из той же широты, что и ликаоны - мельче последних. Среди Canis'ом жару лучше всего переносят, походу, шакалы (даже койот не особо).
Черепа волка и гиеновой собаки в сравнении: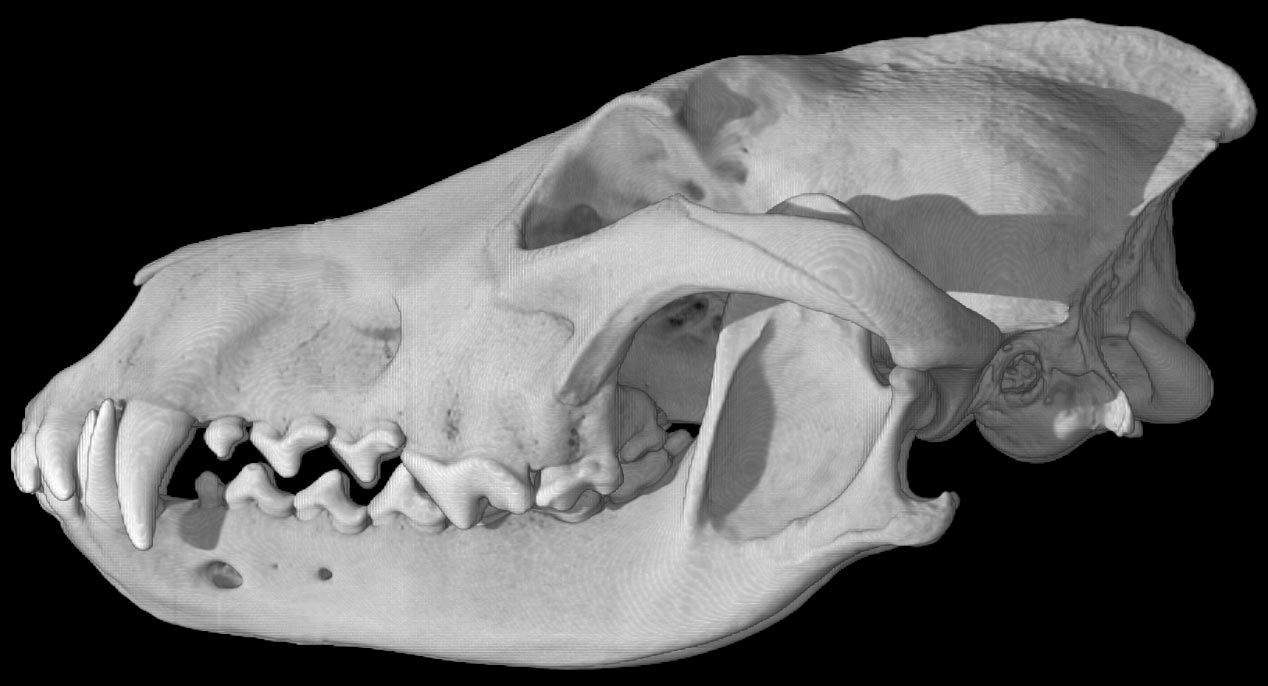
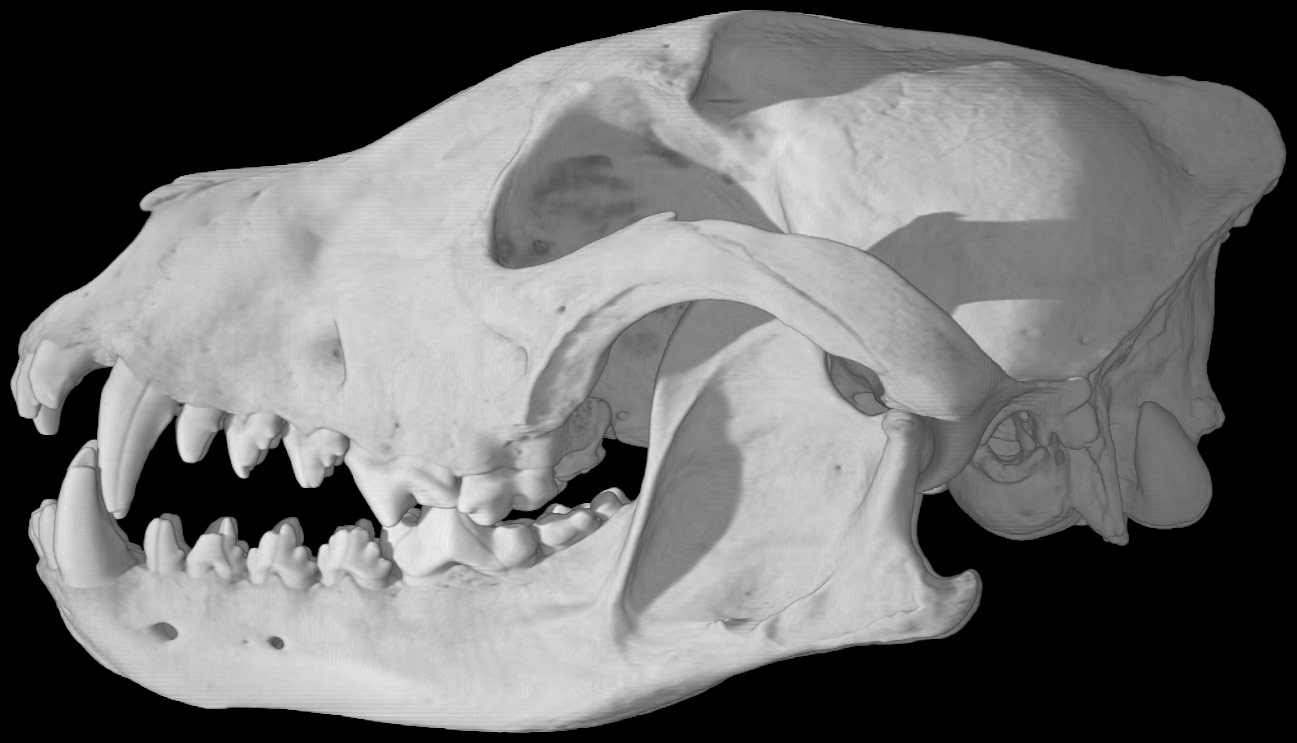
Отредактировано Crazy Zoologist (12 April 2019 17:08:01)
#966 12 April 2019 18:51:22
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Ископаемые псовые (Canidae)

КБ длина в таблице ,общая будет на пару см больше.
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#967 12 April 2019 19:29:54
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Спасибо за конкретный цифры, как всегда. Самый крупный череп из таблицы - 27 см (из Таймыра). Если общая на пару см больше, то получается 29. Череп с линейкой выше по-моему чуть более 30 см. Это очень крупный волк.
#968 12 April 2019 19:51:54
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Ископаемые псовые (Canidae)
Crazy Zoologist :
Спасибо за конкретный цифры, как всегда. Самый крупный череп из таблицы - 27 см (из Таймыра). Если общая на пару см больше, то получается 29. Череп с линейкой выше по-моему чуть более 30 см. Это очень крупный волк.

26 см не больше.
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#969 13 April 2019 13:56:00
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Алекс :
Crazy Zoologist :
Спасибо за конкретный цифры, как всегда. Самый крупный череп из таблицы - 27 см (из Таймыра). Если общая на пару см больше, то получается 29. Череп с линейкой выше по-моему чуть более 30 см. Это очень крупный волк.
https://d.radikal.ru/d06/1904/66/b689a1df5f65t.jpg
26 см не больше.
{kind=link}
Сколько такая особь будет весить? И эти данные о 90 кг волках достоверны?
#970 13 April 2019 14:02:06
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Ископаемые псовые (Canidae)
Crazy Zoologist :
Алекс :
Crazy Zoologist :
Спасибо за конкретный цифры, как всегда. Самый крупный череп из таблицы - 27 см (из Таймыра). Если общая на пару см больше, то получается 29. Череп с линейкой выше по-моему чуть более 30 см. Это очень крупный волк.
https://d.radikal.ru/d06/1904/66/b689a1df5f65t.jpg
26 см не больше.Сколько такая особь будет весить? И эти данные о 90 кг волках достоверны?
Не знаю ,это нужно самому мерить и взвешивать . 90 кг это очень большой волк должен быть. под метр в холке !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#971 13 April 2019 14:38:58
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Средний вес крупнейших попуяци волков составляет чуть менее 50 кг, 70-ые кг волки известны, но 90 кг, думаю, перебор. Как он загонять добычу будет. У волка туловище довольно стройное и грацильное. Дикие псовые они такие. Вот например, если снять с лисицы шкуру, то она будет очень напоинать левретку.
#972 05 May 2019 14:02:09
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
http://www.nhc.ed.ac.uk/index.php?page=493.172.288
Я не понимаю, почему такой простой биоляп кочует из одного источника в другой - так сложно написать правильную четкую зубную форумулу собак. Там не совсем правильная, число моляров перепутано.
#973 15 June 2019 15:17:27
- kimon943
- Любитель животных
- Зарегистрирован: 16 July 2013
- Сообщений: 721
Re: Ископаемые псовые (Canidae)
[img]



[/img]Найдена голова волка в Сибири. Возраст- 40 тыс. лет. Обнаружили еще год назад. Странно, на снимках он на гиеновую собаку похож. Нет? https://phys.org/news/2019-06-perfectly … beria.html
Отредактировано kimon943 (16 June 2019 09:22:03)
Неактивен
#974 16 June 2019 09:12:47
- kimon943
- Любитель животных
- Зарегистрирован: 16 July 2013
- Сообщений: 721
Re: Ископаемые псовые (Canidae)
Все же волк. Пишут, череп 40 см.
https://indicator.ru/photo/2019/06/12/t … ego-volka/
Неактивен
#975 16 June 2019 21:22:23
- George
- Любитель зоологии
- Зарегистрирован: 01 January 2010
- Сообщений: 899
Re: Ископаемые псовые (Canidae)
Crazy Zoologist :
Вот например, если снять с лисицы шкуру, то она будет очень напоминать левретку.
А если кошку бросить в воду, она удивительно похожа на выдру.
Неактивен