Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#926 26 October 2017 13:27:40
- Predator
- Любознательный

- Откуда: Мезозойская эра
- Зарегистрирован: 01 April 2017
- Сообщений: 143
Re: Ископаемые псовые (Canidae)
Плюс ещё пришли люди. Причина это или нет, вызывает много обсуждений. Но конечно же (по моему мнению), это не сыграло решающую роль. Однако нельзя не признать, что они тоже их добивали.
Только потому что динозавры вымерли, мы можем жить.
Неактивен
#927 26 October 2017 14:36:20
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Predator :
Traper :
Насчет главного вопроса - почему изчез дирус. Думаю основная причина в том что он никогда не проявлял интерес к грызунам и мелким животным. Когда началось Великое вымирание древняя американская прерия уже не могла доставлят в таком большом количестве ни живого мяса, ни трупов, крупные медленные травоядные начали исчезат, огромные кости больше не валялись везде по прериям. Дирус имея несколько примитивный мозг не догадался что мыши, белки и полевки - тоже пища. Взял да вымер. Возможно последние проголодавшиеся дирусы умирали печально смотря как какой нибудь суслик или белка рыщут у них под носом

Такая участь постигла и смилодонов, арктодусов и атроксов, но для них оправдание в том что их огромные тела не могли прокормится мелкими млекопитающими, а дирус наверно мог, но не догадался. Исчезли все крупные облигатные хищники, даже пума временно нашла рефиджиум в южноамериканских Анд, прежде чем вернутся. В всей Северной Америке остался только один крупный облигатный хищник, до сих пор незамеченным никем, самый пластичный и самый умный из всех - серый волк. Постепенно серый волк овладел огромные пространства Северного поушария и стал верховным облигатным хищником в всех холодных областей Земли.Безусловно, логика здесь присутствует. Но не могу я поверить, что целый вид вымер только из-за того, что не додумался охотиться на то, что осталось. Даже учитывая тот факт, что он был приспособлен к охоте на больших животных, он мог пытаться охотиться на оставшуюся живность. Но этого было слишком мало даже в случае успеха. Ему нужно было больше пищи, а она исчезла. Безусловно, исчезновение гигантских травоядных играет здесь роль, причём очень важную. Но глобальной причиной вымирания и травоядных и хищников стала перемена климата, исчезновение растительности... В общем, Мать Природа вновь погубила своих детей.
А как же мастодонты и некоторые гомфотерииды которые доживают до голоцена?
#928 26 October 2017 18:12:12
- Predator
- Любознательный
- Откуда: Мезозойская эра
- Зарегистрирован: 01 April 2017
- Сообщений: 143
Re: Ископаемые псовые (Canidae)
Некоторые гигантские травоядные просто вымерли, а некоторые ушли на север.
Только потому что динозавры вымерли, мы можем жить.
Неактивен
#929 26 October 2017 19:14:00
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Не матодонт дальше зоны лесов вроде не удалялся на север.
#930 26 October 2017 19:26:06
- Eugene
- Модератор

- Откуда: Москва
- Зарегистрирован: 12 March 2013
- Сообщений: 5056
Re: Ископаемые псовые (Canidae)
Predator :
Некоторые гигантские травоядные просто вымерли, а некоторые ушли на север.
"Ушел на север, просьба не беспокоить. Ваш гигантский травоядный."
А мы уйдем на север,
А мы уйдем на север
И переждем!
Когда же мы вернемся,
Не будет никого
И даже Лягушонка
И косточек его,
И косточек его...![]()
![]()
![]()
Неактивен
#931 26 October 2017 21:14:31
- Predator
- Любознательный
- Откуда: Мезозойская эра
- Зарегистрирован: 01 April 2017
- Сообщений: 143
Re: Ископаемые псовые (Canidae)
Амфицион :
Не матодонт дальше зоны лесов вроде не удалялся на север.
Ну даже если так, то то, что он ушёл не до конца, не спосло дируса. И не только дируса, но и всех остальных гигантских хищников. Ведь все остальные травоядные исчезли.
Только потому что динозавры вымерли, мы можем жить.
Неактивен
#932 26 October 2017 23:23:26
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Что, и в Евразии тоже?
#933 26 October 2017 23:43:28
- Сorvin
- Модератор

- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5946
Re: Ископаемые псовые (Canidae)
Коллеги, а может, для того, чтобы снизить градус эмоциональности, всем участникам дискуссии стоит признать хотя бы тот факт, что Canis dirus давным-давно вымер (помните финал в сказках про Кощея: «а потом взял – и издох», меня это ещё ребёнком прикалывало). Ничего тут уже не изменишь, это стоит принять как свершившееся и истинную причину вымирания мы вряд ли узнаем, а излагаемые причинно-следственные связи являются лишь нашими гипотезами (причём ничего не решающими), так зачем быть столь агрессивными? Здесь ведь собрались единомышленники и нас, столь скрупулёзно и въедливо увлекающихся вымершими тварями, на самом деле не очень много (сонм детей и юношей, фанатеющих от динозавров, во внимание не беру). Стоит ли клацать друг на друга зубами из-за дохлых зверюг? И, с другой стороны – почему бы не прислушаться к аргументам оппонирующей стороны, если они действительно достойны внимания? Противостоять ради упрямства просто неразумно, нам ведь давно не шешнадцать )), поди. Надо уметь друг друга не только слушать, но и слышать… а то превращаемся в аналог политических ток-шоу, засравших федеральные каналы (слава Дарвину, без кликушества), а наши посты скорее напоминают интеллектуально заострённые стилеты со смазанными ядом завуалированного сарказма стилистически отточенными гранями хорошего русского языка, чем действительно информативные сообщения. Ну понятно, что далеко не дураки или юноши пишут, а серьёзные дядьки и читать посты, даже просто наслаждаясь изощрённым выверенным «логосом», приятно, чёрт возьми!
А вот за глупость надо карать. Как и за хамство.
Братцы, а вот меня всегда больше интересовало возникновение некоей жизненной формы, чем её исчезновение.
На данный момент принято, что C. dirus, скорее всего, потомок североамериканского C. armbrusteri. И что филогенетически лютоволк – сестринский таксон нашего родного серого. Всё! На этом компромисс всех исследователей заканчивается, дальше – взятие Шатоя Жириновским и кого сюда только не приплетали.
Вот кааца, что «дирус» всё же – неарктический эндемик, дивергировавший от «хордовой» линии ещё на уровне волков/койотов. Другая филогенетическая линия «прыснула» в мезамериканский регион (Canis nehringi, C. gezi), а вовсе не наоборот. Но связь с азиатским C. chihliensis для чистоты анализа исключать нельзя, здесь нужна ОЧЕНЬ большая выборка.
Ну, и под занавес – ребят, вот здесь много рассуждали об обрушении североамериканской мегафауны позднего плейстоцена, ориентируясь на «ужасного» волка. Но именно про самого «дируса» на протяжении нескольких страниц ничего ведь и не было… Где сам пёс? Хорошо, пусть дискутирующие знают его метрику с учётом хронопопуляций и гендерных допусков; в курсе относительно темпорально-географического распространения и морфологических характеристик, касаемо как локомоционных аспектов, так и филогенетических связей с другими собачками; осведомлены о фаунистических изменениях в териокомплексах Северной Америки на протяжении плейстоцена – но не увидел «дируса»… C. dirus - вымершая псина, она известна только по костям, а вот ссылок на них не имеется. Коллеги, при всём уважении все вышеизложенные посты являются палеофантазией. Интересной, вариативной, но … наличествующие аргументы не касались конкретно остатков самой собачки. Дискуссия масштабна, однако субъективно по таксону всё очень «размыто».
Может, пора заканчивать?
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#934 27 October 2017 21:15:30
- Gosha01
- Любитель зоологии

- Зарегистрирован: 20 June 2012
- Сообщений: 828
Re: Ископаемые псовые (Canidae)
Может эта статья частично прояснит ситуацию о взаимоотношениях ужасных волков с серыми. Выводы вроде такие: сначала сосуществовали ужасный волк и койот. Койот уменьшился в размерах чтобы избежать конкуренции с ужасным волком. Потом "вклинился" серый волк, но он оставался малочисленным пока не вымер ужасный. Изменения ареалов всех этих видов плохо коррелируют с климатическими данными, поэтому авторы делают вывод что взаимодействия между ними (видимо конкуренция) играли более важную роль.
Biotic responses of canids to the terminal Pleistocene megafauna extinction
MI Pardi, FA Smith - Ecography, 2016 - Wiley Online Library
Our findings indicate that interactions among survivors within the canid community dramatically changed after the end Pleistocene event. Overall, species changed their spatial distribution, but did not track climatic niche space (Fig. 3-6), which suggests a role for changing biotic interactions within the community. Additionally, we identify a net increase in niche partitioning among surviving canids after the Pleistocene that is independent of climate (Fig. 7), following the extirpation of their largest competitors.
The differing biogeographic histories of the three largest canids in our study system may help explain the patterns we observe from our niche modeling approach. Unlike Canis dirus and Canis latrans, which are long time endemics in North America with a history of interaction, Canis lupus was a relative newcomer to the scene in the Late Pleistocene (Merriam 1912, Dundas 1999). Canis lupus occurred in low densities relative to C. dirus during the Late Pleistocene, and their numbers increase following the extinction of C. dirus (Merriam 1912). Canis latrans was larger during the Pleistocene and would have been similar in size to C. lupus when it initially expanded into North America (Meachen and Samuels 2012, Meachen et al. 2014), so these two species may have initially been competing more directly for resources. The increased abundance of a competitor with a similar dietary niche may have promoted partitioning of resources by C. latrans and induced a shift in its niche during the Holocene. This hypothesis is consistent with other studies that identify body size reduction and morphological changes towards omnivory in coyotes in the Holocene (Meachen and Samuels 2012, Meachen et al. 2014). These are physical responses that might be expected in a species that is drastically changing its niche and ecological function.
In this study we asked whether the niches of endemic North American canids were conserved following the end Pleistocene event. We find that niches were not conserved for any of the species we analyzed.
Неактивен
#935 28 October 2017 23:24:39
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5946
Re: Ископаемые псовые (Canidae)
Да, тёзка, спасибо - в самом деле итогово и толково))). Лучше бы, конечно, перевод, ведь не все англо-буржуйский знают.
Коллеги, давайте здесь пока многоточие поставим и дух переведём хучь на недельку...
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#936 12 November 2017 13:10:25
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
New Fossil Fox Discovered in South Africa, Co-Existed with Australopithecus
http://www.sci-news.com/paleontology/article00840.html
#937 12 November 2017 22:31:37
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
https://www.researchgate.net/figure/228 … -FAM-61746
Эндокраны разных псовых.
#938 14 November 2017 12:43:46
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
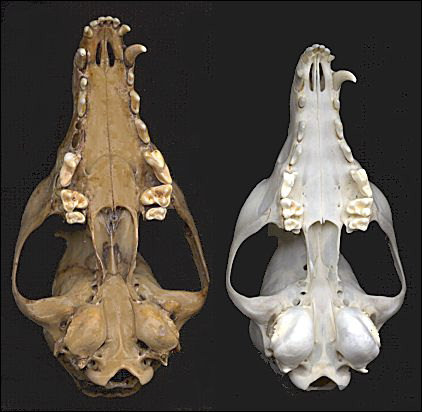
Ископаемый череп американского корсака и череп современной американской лисицы Vulpes macrotis.
https://www.utep.edu/leb/pleistnm/taxaMamm/Vulpes.htm
#939 03 December 2017 00:22:02
- Wolf
- Без пяти минут зоолог

- Зарегистрирован: 13 January 2009
- Сообщений: 2583
Re: Ископаемые псовые (Canidae)
Очень наглядно - филогенетическое дерево псовых:


Отредактировано Wolf (03 December 2017 00:23:26)
Per aspera ad astra
Неактивен
#940 03 December 2017 00:31:15
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Сункахетанка - прикольое название. Что означает? Почему ксеноцин ближе к дхолям, а не гиеновым собакам? Это же их наиболее вероятный предок. Этого Cuon'а куда только не суют, как я посмотрю. В любом случае они очень дальние родственники. Посуди сам: у обоих видов прогрессивная окраска, но у ликаона она разноцветная, а у дхоля одноцветная. Куон в процессе эволюции потерял ненужные моляры, ликаон ненужные пальцы. Соц. организация схожа, так как это псовые "нового образца".
#941 03 December 2017 01:18:31
- Wolf
- Без пяти минут зоолог
- Зарегистрирован: 13 January 2009
- Сообщений: 2583
Re: Ископаемые псовые (Canidae)
Почему ксеноцин ближе к дхолям, а не гиеновым собакам?
По-моему, на дереве это и не отображено..
Я сейчас больше по геспероционам и борофагинам метаюсь, но насколько я помню, считается, что Xenocyon - наиболее вероятный предок обоих дхоля и гиеновой собаки. Раньше к ним еще кустарниковую собаку запихивали на основании похожего строения нижнего хищнического зуба, но сейчас последняя вместе с другими южноамериканскими псовыми.
Это филогеническое дерево морфологи составляли, что в принципе оправданно для геспероционин и борофагин, но в отношении современных псовых было бы интереснее посмотреть, что думают генетики. Так что привожу табличку сделанную на основе выводов работы Genome sequence, comparative analysis and haplotype structure of the domestic dog (Toh et al. 2012):
Тут гиеновую собаку и дхоля не ответвляют отдельно, но все же соблюдают их родственность. Только прикол в том, что получается, что таксон Canis является парафилическим таксоном, потому что полосатый и чепрачный шакалы менее родственны прочим Canis, чем те же самые Lycaon и Cuon. Поэтому нужно либо гиеновых собак и дхоля включить в Canis, либо чепрачного и полосатого шакала объединить в один род..
Отредактировано Wolf (03 December 2017 01:35:07)
Per aspera ad astra
Неактивен
#942 03 December 2017 01:58:17
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Верно рассуждаешь. Положение (нынешнее) в систематике этих двух шакалов меня сильно удивляет. Насчет ликаона посмотри это Hartstone-Rose, A.; Werdelin, L.; De Ruiter, D. J.; Berger, L. R.; Churchill, S. E. (2010). "The Plio-Pleistocene Ancestor of Wild Dogs, Lycaon sekowei n. sp". Journal of Paleontology. Он известен из позднего плиоцена, значит по времени с ксеноционом пересекался. Тот вроде тоже не известен раннее чем из позднего плиоцена неизвестен. А L. lycanoides более поздний, чем этот ликаон.
#943 04 December 2017 19:17:50
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5946
Re: Ископаемые псовые (Canidae)
Денис, спасибо! Подозреваю, откуда был посыл, но за приватный анализ низкий поклон.
Только не спеши, дружище. Откладывай все эти вопросы на "потом" - ты ещё к ним вернёшься, более вооружённый.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#944 08 December 2017 23:46:50
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
3-4 млн. лет назад, согласно диаграмме, отделилась ветвь гиеновых собак. За это время появляются и эволюционируют все новые виды рода Canis, а род Lycaon отпочковывается от Canis намного позже. Как так? Ведь 3-4 и даже 2 млн. лет назад предки гиеновых собак не выделились из Canis.
#945 13 March 2018 23:59:55
- Crazy Zoologist
- Гость
Re: Ископаемые псовые (Canidae)
Нестор, ты как-то давно писал о дхолях на Кавказе. Откуда инфа? Я нигде не находил до сих пор. Даже в журнале Mammalian Species касательно плейстоценовых находок дхолей указана лишь Австрия. И вообще посвящено этому всего пару предложений.
#946 26 May 2018 21:49:55
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5946
Re: Ископаемые псовые (Canidae)
Crazy Zoologist :
как-то давно писал о дхолях на Кавказе. Откуда инфа?
Поиск занял ровно минуту. Взгляни работы Барышникова и Оводова. Куонподобные волки на Урале, в Сибире и Приморье тоже наличествовали.
Странно - если в Европе найдены, то на Кавказе их не было, сам понимаешь.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#947 18 September 2018 16:06:29
- George
- Любитель зоологии
- Зарегистрирован: 01 January 2010
- Сообщений: 899
Re: Ископаемые псовые (Canidae)
В Канаде нашли мумию волчонка возрастом 50 тыс. лет.
https://news.mail.ru/society/34761768/?frommail=1
Неактивен
#948 30 December 2018 11:29:40
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5946
Re: Ископаемые псовые (Canidae)
Коллеги, моё искреннее почтение и всех с Новогодьем!
За десятилетие присутствия на Форуме почти не узрел толковых постов по дохлым собачкам, хотя иногда «высверки» бывали. Ввиду собственного пробела по этой группе как-то пытался менторски стимулировать пару способных и интересующихся вымершими псовыми форумчан приобщить к анализу, но уже более года, как «выхлопа» НЕТ. Посему «старая гвардия» решила взять инициативу в собственные руки. Итогом является серия нижеизложенных постов, иллюстрирующих систематику канид в современном воззрении.
Сразу скажу, что изначальной задачей было первичное ознакомление с имеющимся информационным массивом, а в итоге - лишь «пробег», конечной целью которого являлось выявление основных морфологических тенденций в семействе. Группа интересная и в ней надо «сидеть», тут очень много нюансов.
Посты «суховаты» в освещении морфологических отличий и сравнительного анализа между таксонами – не стал текстово перегружать, а кто желает узнать подробнее – пжалте в «личку», имеющейся информацией с удовольствием поделюсь. Также не стал громоздить осмотры современных таксонов – к примеру, эволюционная история хотя бы енотовидной собаки заслуживает отдельного обзора как минимум, так как она начинается ещё в миоцене, ареал охватывал всю Палеарктику (Европу и Африку в том числе), а до наших дней из всего разнообразия дожил всего один дальневосточный вид. То же самое касается Xenocyon – такое впечатление, что эта форма (возможно, породившая куонов и ликаонов) была в плиоцен-раннем плейстоцене столь же морфовариативна, как Canis. Впрочем, как и Eucyon – по большому счёту, эта группа в систематике Canidae вообще отдельной подтрибы заслуживает.
Диагнозы также «кастрированы» - согласитесь, кому интересно кроме пары-тройки интересующихся парней читать о морфологических нюансах? Вот только нередко именно на их основе и вычленяется новый таксон.
Признаю, что придерживался «академичности», но иногда позволял приватные ремарки в плане собственного мнения - это в тексте видно. Также признаю некоторую субъективность с моей стороны, так как основное внимание уделено именно ископаемым таксонам. Но ведь без анализа таковых невозможно адекватно рассуждать об эволюции современных, не так ли? При рассмотрении таксонов в ОБСУЖДЕНИИ – собственные приватные мыслишки.
Скажу, что даже при беглом осмотре в эволюционной истории канид имеется гораздо больше «белых пятен», чем у кошачьих. И очень негативен тот вопрос, что 2/3 эволюции псовых происходило на территории Северной Америки.
Огромное спасибо Роману Станиславовичу, как всегда, альтруистично присоединившемуся к этому проекту.
Также не исключено, что при дальнейшем изучении точки зрения могут несколько измениться, поэтому в анализе радикальность собственного мнения старался минимизировать.
По сути, Canidae выделяются от других Carnivora в особенностях морфологии слухового региона, наличии задних дополнительных бугорков на премолярах; редукции парастиля на М1 и исчезновении M3.
Большинство базальных характеристик присущи всем представителям Canidae (за исключением амфицидонид, которые морфологически более арктоидны). Систематически на уровне семейства в консервативном воззрении наличествует дифференция на три подсемейства и в настоящее время эта точка зрения формирует основные рамки филогенеза канид. Как бы то ни было, несмотря на их высокое специфическое разнообразие, каниды демонстрируют ограниченный диапазон морфологических моделей на протяжении всей их истории – схожие дентальные структуры появляются у разных групп независимо. Таким образом, филогенетические анализы часто оказываются разочаровывающими.
Также возможно, что в подсемействах Hesperocyoninae и Borophaginae присутствуют миоценовые «аппендиксные» таксоны, иллюстрирующие их полифилетичность. К примеру, подсемейство Hesperocyoninae, очерченное Тедфордом (1978), строго не определено – Тедфорд просто объединил в одну группу самых примитивных канид, объединённых морфологическими характеристиками, которые по большей части примитивны для всех псовых.
Из базальных форм внимания заслуживают следующие:
Canidae incertae cedis
LYCOPHOCYON Tomiya, 2011
Lycophocyon hutchisoni (UCMP 85202), голотип
ТИПОВОЙ ВИД: Lycophocyon hutchisoni
РАСПРОСТРАНЕНИЕ: Калифорния, дюшений (средний эоцен)
ДИАГНОЗ: отличается от каноидных не полностью окостеневшим энтотимпаником, прочно приросшим к базикраниуму, от других хищников - широкой и плоской петрозалью, морфологией М1, третьих премоляров и пр.
ОБСУЖДЕНИЕ: классифицируется как каниморфный хищник на основе результатов кладистического анализа, хотя принадлежность к определённому семейству в настоящее время с нынешним знанием форм и базальной филогении хищных млекопитающих остаётся под вопросом.
PROHESPEROCYON Wang, 1994
Prohesperocyon wilsoni (TMM 40504-126), голотип
ТИПОВОЙ ВИД: Hesperocyon wilsoni Gustafson, 1986.
РАСПРОСТРАНЕНИЕ: юго-западный Техас, чадроний (поздний эоцен).
ДИАГНОЗ: обладает типичной базовой краниодентальной морфологией канид, отличной от миацид: наличие полностью окостеневшего энтотимпаника, медиальное расширение петрозали, наличие задних бугорков на р3, уменьшение внутреннего цингулума и парастиля на М1, отсутствие М3 и пр.
ОБСУЖДЕНИЕ: на сегодняшний день является самым примитивным ископаемым псовым, а его общая морфология довольно близка к североамериканскому Procynodictis vulpiceps (=Miacis gracilis) . Густафсон (Gustafson, 1986) указал, что P. wilsoni уже является истинным канидом, что подтверждает наличие полностью окостеневшего энтотимпаника наряду с другими основными чертами.
К слову, известны находки базальных канид из Монголии, но некоторыми исследователями они, вроде бы, относятся к амфиционидам. В любом случае многие ископаемые находки лишь подтверждают, как мы мало знаем о том эволюционном разнообразии, которого никогда уже не увидим.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#949 30 December 2018 11:39:10
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5946
Re: Ископаемые псовые (Canidae)
Подсемейство HESPEROCYONINAE Tedford, 1978
ДИАГНОЗ: более прогрессивные, чем базальный Prohesperocyon и миациды. Характеризуются мозаичной морфологией слуховой буллы, медиально расположенным протоконом на P4, удлинёнными хищнические зубы, а также редуцированным/исчезнувшим передним цингулумом на M1. Также представители Hesperocyoninae имеют заострённый талонид на m1 (за исключением Osbornodon) в отличие от сниженных талонидов у Borophaginae и Caninae. Кроме того, отсутствует метаконуль на M1, который часто наличествует у борофагин и канин.
РАСПРОСТРАНЕНИЕ: поздний эоцен - средний миоцен Северной Америки.
HESPEROCYON Scott, 1890
ТИПОВОЙ ВИД: Canis gregarius Cope, 1873.
ДИАГНОЗ: уменьшенный и медиально позиционирующийся протокон на P4; увеличенный внутренний цингулум на M1; тригонид и талонид на m1 почти равной ширины. Кроме того, отличается от Prohesperocyon метаконом на М2 почти такого же размера, как паракон и протоконид и метаконидом на m2.
РАСПРОСТРАНЕНИЕ: ранний арикарий Северной Америки (30-25 млн) – граница олигоцена и миоцена.
ОБСУЖДЕНИЕ: род претерпел длинную и довольно любопытную историю номенклатурных изменений. Лейди (Leidy, 1856) сначала кратко описал фрагмент левой нижнечелюстной ветви с p4- m1 из Небраски (ANSP 11072), назвав его Amphicyon (?) gracilis, не подозревая, что это название уже занято европейским таксоном A. gracilis Pomel, 1847. Впоследствии его именовали Canis gregarius, Galecynus, Cynodictis, пока Скотт (Scott, 1890) не предложил новый род Hesperocyon с формальным обозначением типового вида.
Возможно, что Hesperocyon на самом деле – парафилетичен. Уонг (Wang, 1957) провёл колоссальное исследование - помимо изучения морфологических аспектов, выявив эволюционные аналогии, поставив эволюционную «метку» и многоточие в таком вопросе, что Hesperocyon может являться предком как для борофагин, так и для канин.
ПРИМЕЧАНИЕ: H. paterculus по Уонгу не признаётся валидным таксоном, а H. lippincottianus рассматривается им как как возможный дафоенид.
Череп и мандибула Hesperocyon gregarious (USNM 437888)
Есть ещё две формы геспероциона, которые взяты в «кавычки» ввиду того, что они могут к данному роду на самом деле и не принадлежать:
“Hesperocyon” pavidus
ГОЛОТИП: LACM 466, разрушенная передняя половина черепа с зубным рядом из раннего арикария Северной Америки
ОБСУЖДЕНИЕ: малоразмерная форма геспероциона из нескольких местонахождений северо-восточного Колорадо (пять челюстных фрагментов) была описана как Pseudocynodictis sp. (Galbreath, 1953), но от предоставления им формального имени исследователь воздержалась, хотя указано на достаточное разделение от H. gregarious. Уонг (Wang, 1990) признаёт их как новый вид (вместе с более полным материалом из Вайоминга) по ряду морфометрических и геохронологических причин, считая некоторые образцы по некоторым признакам базальными представителями борофагин.
“Hesperocyon” coloradensis 
Череп и мандибула "Hesperocyon" coloradensis (KUVP 85067), голотип
ГОЛОТИП: KUVP 85067 - фрагментарные череп и нижняя челюсть с зубным рядом, фрагментарный посткраниальный скелет.
ДИАГНОЗ: нижние челюсти глубже и массивнее, слуховая булла крупнее из-за увеличения энтотимпаника, М1 трансверсально более удлинён.
ОБСУЖДЕНИЕ: В 1873 году был описан Canis (Hesperocyon) lippincottianus и с тех пор каждый экземпляр «боролся» за свою валидность. Путаница, похоже, связана со статусом голотипа. Одно время его консолидировали с Daphoenus minimus, затем дискутировали о морфологии талонида и о возможном половом диморфизме (ввиду крупного размера). В итоге Уонг (Wang, 1990) вывел его как отдельную форму на основании некоторых дентальных особенностей (образец AMNH 5327a имеет очень крупный энтоконид, который занимает более трети талонида и позиционируется как заострённый выступ, в отличие от Hesperocyon, энтоконид которого обычно является не более, чем редуцированным бугорком, спроецированным лингвально по отношению к гипокониду). Данная особенность наблюдается и у других крупноразмерных образцов геспероционов. К тому же размеры этой формы весьма превышают ожидаемую разницу в половом диморфизме между самцами и самками H. gregarius – т. е. это явно другая «морфа», что наглядно наблюдается и в морфометрии дентальных элементов.
Ещё один небольшой образец фрагментарных челюстей из Колорадо (упоминается как Pseudocynodictis sp.) некогда был включен в гиподигму Cormocyon pavidus. Однако талонидная структура этого образца скорее представляет собой Hesperocyon, а подобная морфология несовместима с филогенетической гипотезой ранних борофагов, которая подразумевает, что если сестринская связь между Borophaginae и Caninae правильна, то талонид должен был развиться во время ореллания, поскольку он уже имеется у раннего Leptocyon. Таким образом, небольшая форма из ореллания Колорадо может представлять собой отдельный вид Hesperocyon.
MESOCYON Scott, 1890
ТИПОВОЙ ВИД: Mesocyon coryphaeus (Cope, 1879).
ДИАГНОЗ:отличается от Hesperocyon и Paraenhydrocyon позицией и морфологией парокципитального выступа и общим видом М2.
РАСПРОСТРАНЕНИЕ: арикарий Северной Америки.
ОБСУЖДЕНИЕ: ряд форм объединён в единый таксон на основе сходной морфологии нижних хищнических зубов. Известны Temnocyon и Hypotemnodon (H. coryphaeus) , находящиеся в таксономической «обойме» Mesocyon. В настоящее время является сборным таксоном для среднеразмерных канид позднего олигоцена Северной америки. Также, очевидно, парафилетичен.
Mesocyon coryphaeus
[url=https://radikal.ru] [/url
[/url
Череп Mesocyon coryphaeus, AMNH 6859 (голотип)
Принят Коупом (Cope, 1879) как архетип позднеолигоценовых канид Северной Америки со множеством вариативных форм (к примеру, подвид M. josephi secundus (YPM 10063) с наличием Р2, хотя это, наверное, ювенильная особь). В настоящее время известно около десятка образцов, приписываемых данному таксону, в то же время до сих пор ведутся дискуссии о принадлежности некоторых к Cynodesmus.
“Mesocyon” temnodon 
Череп и мандибула "Mesocyon" temnodon, AM 63367
Обладает развитыми задними дополнительными и цингулумными выступами на всех верхних и нижних премолярах - этого не наблюдается ни в других геспероционинах, ни в борофагинах. Из других образцов, отнесённых к этому таксону, интересно следующее:
Несмотря на то, что M. temnodon и Paraenhydrocyon josephi - оба примитивных представителя геспероционов и «их очень трудно отличить только по морфологии зубов» (Тедфорд), часть образцов из локации Шарпс (Sharps Formation), ранее считаемых Paraenhydrocyon robustus, следует отнести к Mesocyon по следующим признакам: наличие выраженного заднего дополнительного бугорка на нижних премолярах, относительно большие метакониды на m1 и m2, довольно расширенный энтоконид на талониде m1 и пр.
Mesocyon brachyops 
Череп Mesocyon brachyops, UCMP 1692 (голотип)
Вроде бы его объединили с M. baileyi, дифференцировав имеющиеся образцы по М1, но этот вопрос дискутивен.
На самом деле есть ещё полдюжины образцов из сходных фаунокомплексов, которые синонимически «притянуты за уши».
CYNODESMUS Scott, 1893
ТИПОВОЙ ВИД: Cynodesmus thooides
ДИАГНОЗ: Cynodesmus отличается от других геспероционов увеличенным парокципитальным бугорком с задним выступом, а также хорошо развитым М2. Среди геспероционов отличается менее массивными премолярами и несколько укороченным m1 с развитым метаконидом - такое не наблюдается у Sunkahetanka, Philotrox и Enhydrocyon.
РАСПРОСТРАНЕНИЕ: поздний арикарий – ранний уитний Северной Америки (примерно 3,3 млн).
Cynodesmus thooides 
Череп и мандибула Cynodesmus thooides, YPM-PU 12730 (голотип)
ГОЛОТИП: YPM-PU 10412, череп с почти полным верхним зубным рядом, кроме правого 11-2, P1 и М2, а также нижняя челюсть.
ДИАГНОЗ: отличается от сестринского таксона C. martini меньшим размером, не столь массивными зубами, утоньшенным рострумом и пр.
ОБСУЖДЕНИЕ: дивергенция Cynodesmus от Hesperocyon несколько затруднительна для полновесного анализа ввиду микродонтии P4-M1 (присущей также Mesocion и Sunkahetanka): P4 относительно M1 начинает удлиняться только в родах Enhydroconon и Osbornonon.
Исследование черепа Mesocyon drummondanus (CMNH 792) показало, что он, очевидно, принадлежит C. thooides согласно более меньшей, но массивной слуховой булле, хотя и отличается от типового вида некоторыми дентальными особенностями.
Макдональдом (1970) на основании частичной нижней челюсти с почти полными зубными рядами (LACM 17003) выводился новый вид Brachyrhynchocyon douglasi, удлинёнными премолярами и увеличенным задним бугорком на р4 схожий с Osbornodon. Однако нередуцированная ширина его предкоренных зубов, похоже, исключает эту возможность и в большинстве других случаев этот образец премолярным комплексом очень схож с C. thooides, за исключением немного меньшего m1.
Cynodesmus martini
ГОЛОТИП: UNSM 25796, почти полный череп и нижняя челюсть с левыми I2-3 и P2-M2, правыми I3 и P2-M2, и левый c1 и p4-m3.
РАСПРОСТРАНЕНИЕ: ранний арикарий Небраски.
ДИАГНОЗ: обладает рядом продвинутых признаков, отличающих его от C. thooides: гораздо более крупным и массивным черепом, более широкой носовой частью и вытянутой затылочной с высоким сагиттальным гребнем, а также исчезновением Р1.
ОБСУЖДЕНИЕ: достигает размера и пропорций Enhydrocyon, а также сходными дентокраниальными характеристиками, однако большинство сходств между C. martini и Enhydrocyon рассматриваются как независимые «попытки» C. martini в развитии гиперкарниворных адаптаций.
SUNKAHETANKA Macdonald, 1963
Голотип Sunkahetanka geringensis (UNSM 1092)
ТИПОВОЙ ВИД: Sunkahetanka geringensis Macdonald, 1963.
ГОЛОТИП: UNSM 1092, полный череп и нижняя челюсть с почти целым зубным и посткраниальным скелетом.
РАСПРОСТРАНЕНИЕ: ранний миоцен Северной Америки
ОБСУЖДЕНИЕ: Макдональд (Macdonald, 1963) предложил новый род Sunkahetanka для Mesocyon geringensis, который был «производным от линии мезоционов». Барбур и Шульц (Barbour and Schultz, 1935) ещё раньше перечислили ряд отличительных признаков M. geringensis от типового вида M. coryphaeus.
Большинство этих характеристик демонстрируют собой общую тенденцию у Sunkahetanka к более массивному черепу и крупному зубному ряду по сравнению с Mesocyon и Cynodesmus. Sunkahetanka демонстрирует определённые признаки развития зубов, присущие Enhydrocyon (Philotrox в меньшей степени), но укороченная задняя часть черепа, низкий сагиттальный гребень и мелкая мандибула подтверждает, что S. geringensis всё ещё не является истинным Enhydrocyon. К последнему принадлежит другой экземпляр, первоначально именованный как S. pahinsintewakpa. Третий вид, S. sheffleri (LACM 17076), теперь синонимизирован с C. thooides и представляет собой популяцию, которая, наверное, является предком собствено S. geringensis.
PHILOTROX Merriam, 1906
Череп и нижняя челюсть Philotrox condoni (UCMP 76712)
ТИПОВОЙ ВИД: Philotrox condoni
ГОЛОТИП: UCMP 89, задняя часть черепа и фрагмент левой ветви нижней челюсти с i3-m3
РАСПРОСТРАНЕНИЕ: ранний арикарий Северной Америки
ДИАГНОЗ: в отличие от более примитивного S. geringensis, Philotrox имеет такие производные признаки, как удлинённая посторбитальная область черепа, глубокая масеттерная ямка, высокий сагиттальный гребень, укрупнённая булла, более широкие премоляры и пр.
ОБСУЖДЕНИЕ: в настоящее время Philotrox почему-то не привлекает большого внимания исследователей, хотя обладает многими мозаичными признаками с Enhydrocyon и Sunkahetanka. Выглядит как переходная форма между таковыми, но редкость находок не позволяет сделать однозначные выводы. В то же время обладает сходными признаками с «короткомордыми» псовыми.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен
#950 30 December 2018 12:49:54
- Сorvin
- Модератор
- Откуда: Двор Хаоса
- Зарегистрирован: 28 December 2009
- Сообщений: 5946
Re: Ископаемые псовые (Canidae)
OSBORNODON Wang, 1990

ТИПОВОЙ ВИД: Osbornodon fricki
ДИАГНОЗ: даже в самых примитивных формах Osbornodon имеет 3 синапоморфии, отличающих его от остальных геспероционов: вентральная направленность парокципитального выступа; более квадратные верхние моляры и увеличенные вторые коренные зубы. Впоследствии виды Osbornodon постепенно приобретают дополнительные производные символы: относительно удлинённый рострум; куполообразность лобной области, расширяющийся назад парокципитальный выступ и увеличенный p4.
ОБСУЖДЕНИЕ: несмотря на то, что виды Osbornodon легко отличить от других родов Hesperocyoninae, примитивные формы морфологически гораздо менее отчётливы, особенно при диагностике изолированных зубов. Осборнодон параллелен подсемейству Caninae в некоторых особенностях черепа и зубов: тонкий рострум, удлинённые премоляры и пр.. Талонидный бассейн принципиально отличается от такового у Borophaginae и Caninae, у которых отсутствует поперечный гребень между гипоконидом и энтоконидом. Талонидный бассейн у Osbornodon окружен гребневидным энтоконидом на медиальной стороне, а не выступом, как у борофагов и канин.
Osbornodon fricki 
Образец Osbornodon fricki (FAM 27363), голотип
ГОЛОТИП: FAM 27363 - череп с почти полным зубным набором и частичная посткраниаль.
ОБСУЖДЕНИЕ: демонстрирует прогрессивные зубные характеристики, свойственные гиперкарниворным хищникам - удлинённый Р4, массивный p4, уменьшенный метаконид на m1. С другой стороны, O. fricki обладает зубными особенностями, характерными для мезокарниворных канид: квадратный М1, относительно увеличенный М2 и m2, а также талонид на нижних коренных зубах. 
Osbornodon renjiei
ГОЛОТИП: RAM 63316 - верхнечелюстные и нижнечелюстные фрагменты с клыками и премоляр-молярными рядами.
ОБСУЖДЕНИЕ: как отдельный таксон в значительной степени признан по морфологии зубов.
Osbornodon sesnoni
ГОЛОТИП: LACM 17039 - частичный череп и нижняя челюсть, посткраниальные фрагменты.
ОБСУЖДЕНИЕ: Макдональд (Macdonald, 1970) выводил род Brachyrhynchocyon с двумя видами B. sesnoni и B. douglasi. Однако не сформулировал морфологическую основу этих присвоений, на основании чего изученный им материл был распределён в амфициониды и каниды - Cynodesmus thooides и данная форма как валидный таксон.
Osbornodon iamonensis
ГОЛОТИП: USNM 8836 (ранее FGS 5082) - правый верхнечелюстной фрагмент с P4-M2.
ОБСУЖДЕНИЕ: Симпсон (Simpson, 1932) установил вид Cynodesmus nobilis по восьми образцам из одной локации и указал, что он «очень схож с Mesocyon iamonensis» (на основе крупного М2). Вообще, таксономическая история этой формы довольно забавная и длинная – к примеру, её выделяли в отдельный вид Paradaphaenus tropicalis на основе большого гипоконида на m1 (White, 1942), причём диагноз Уайта почти полностью противоположен Симпсону.
В свете довольно гомологичной черепной морфологии в образцах Флориды и Небраски Уонгом (Wang, 1990) они считаются конспецифичными и сведены в общую форму. O. iamonensis является филогенетически первым прогрессивным представителем рода и многие производные символы можно увидеть в завышенном лобном синусе, высоком сагиттальном гребне, расширенном парокциптальном выступе, увеличенном р4, уменьшенном Р3, удлинённом Р4 и увеличенном М2.
Osbornodon brachypus
ГОЛОТИП: AMNH 8140 - череп и нижние челюсти плюс большая часть посткраниального скелета.
ОБСУЖДЕНИЕ: Коуп (Cope, 1883) сравнивал описанного им Canis brachypus с современным койотом (C. latrans) и интерпретировал его как «самый старый вид нынешних псовых, известный мне из американских форм». Ему вторили Скотт (Scott, 1895) и Петерсон (Peterson, 1910), использовавшие его в качестве примера вида, близкого к «истинной» линии канид, в отличие от более аберрантного Daphoenodon superbus. В то же время большинство синапоморфий, объединяющих O. brachypus и O. fricki, явно выделяют его из родословной канин.
PARAENHYDROCYON Wang, 1990
Paraenhydrocyon wallovianus (AMNH 81086)
ТИПОВОЙ ВИД: Paraenhydrocyon wallovianus
РАСПРОСТРАНЕНИЕ: ранний - поздний арикарий Северной Америки
ДИАГНОЗ: представители Paraenhydrocyon отличаются от других родов геспероционин высокими и остроконечными основными бугорками и редуцированными вспомогательными на премолярах, а также редуцированным тригонидным бассейном на M1 (для приёма гипоконида m1).
ОБСУЖДЕНИЕ: Paraenhydrocyon разделяет тенденцию Enhydrocyon в развитии гиперкарниворности хищнических зубов. Однако общая форма премоляров и общие пропорции черепов настолько различны между Paraenhydrocyon и Enhydrocyon, что они явно принадлежат к разным эволюционным линиям. Это также подтверждается различной топологией парокципитальных выступов – в целом, уникальная комбинация канидных и амфицидонодоподобных признаков делает Paraenhydrocyon уникальным таконом, весьма отличающимся от остальных геспероционов.
Paraenhydrocyon wallovianus
ГОЛОТИП: AMNH 6858 - измельченная ростральная часть черепа с сохранившимися зубными рядами.
ОБСУЖДЕНИЕ: P wallovianus является самым крупным и, насколько пока известно, терминальным членом рода. Наблюдается прогрессивный эволюционный тренд в морфологии m1-m2 (линия P. robustus - P. wallovianus). Однако не исключено, что они являются сестринскими таксонами. Интересно замечание Стивенса (Stevens, 1991), изучавшего его посткраниум, что P. wallovianus не являлся ярко выраженным курсориальным хищником, а вполне мог даже изредка взбираться на деревья.
Paraenhydrocyon josephi 
Paraenhydrocyon josephi (YPM 12702)
ГОЛОТИП: AMNH 6878 - передняя половина черепа с полными верхними зубами, кроме клыков и резцов.
ОБСУЖДЕНИЕ: довольно вариативный вид, так как образцы из раннего арикария (конец олигоцена) демонстрируют размерный «разброс» слуховой буллы, развитие вспомогательных премолярных бугорков, а также размер и форму верхних коренных зубов. Образцы из позднего арикария (ранний миоцен), представленные хорошо сохранившимися черепами, уже показывают небольшое увеличение размера и случайные вариации, которые похоже, не ведут в каком-либо конкретном эволюционном направлении.
Paraenhydrocyon robustus
ГОЛОТИП: AMNH 12884 – верхнечлюстные фрагменты с почти полными зубными рядами.
ОБСУЖДЕНИЕ: первоначально именовался как Mesocyon hortulirosae (Schlaikjer, 1935), но впоследствии образцы согласно более производной морфологии коренных зубов отнесены Макдональдом (Macdonald, 1963) к отдельному таксону, позже консолидированных с P. robustus,
CAEDOCYON Wang, 1990
Caedocyon tedfordi (10251), голотип
ТИПОВОЙ ВИД: Caedocyon tedfordi
ГОЛОТИП: UW 10251 - ростральная часть черепа с левыми I3-M1 и правыми C1-M2.
ОБСУЖДЕНИЕ: филогенетические связи C. tedfordi озадачивают главным образом из-за неполного знания черепной и зубной анатомии и отсутствия промежуточных форм. Без этого знания, по большому счёту, даже его принадлежность к канидам, как таковым – под вопросом. Однако его чрезвычайно редуцированный М2 свидетельствует о его маловероятной принадлежности к амфиционидам, так как зубы Caedocyon скорее всё же каниморфны.
Принадлежность к канидам выводится по гомологичным признакам кладистики, которые сближают Caedocyon с Paranenhydrocyon – по дентальным и черепным признакам. Уонг признаёт, что нынешний предварительный морфологический паритет Caedocyon с Paranenhydrocyon лишь не что иное, как предположение, ожидающее проверки по более полным материалам.
ECTOPOCYNUS Wang, 1990
Нижние челюсти Ectopocynus (голотипы): А: Е. antiques (FAM 54090); B: E. intermedius (FAM 27580); C: E simplicidens (FAM 25431)
ТИПОВОЙ ВИД: Ectopocynus simplicidens
РАСПРОСТРАНЕНИЕ: поздний арикарий Северной Америки
ДИАГНОЗ: имеет короткие, тупые и прочные нижние премоляры с чрезвычайно уменьшенными/отсутствующими дополнительными выступами и цингулумными бугорками. За исключением одного образца, все нижние первые премоляры у всех утрачены. Метакониды m1-2 редуцированы даже в самых примитивных формах, в отличие от других линий геспероционов, у которых метакониды уменьшаются только в крупноразмерных прогрессивных формах.
ОБСУЖДЕНИЕ: материал наситывает только шесть экземпляров, представляющих три вида этой чрезвычайно редкой линии геспероционов в течение геологического интервала примерно около 12 миллионов лет от уитния до среднего хемингфордия (? !!!).
Филогенетическая связь Ectopocynus неизвестна. Отсутствие знаний о его краниальной анатомии затрудняет её подтверждение родство к канидами. Но на самом деле, в сравнении с хемингфордийским Aelurocyon имеются отличия в более сниженном венечном отростке, маловыраженном внутреннем гребнем углового выступа, крупном М2, удлинённом m2 и наличии m3. И здесь разумно предположение Уонга (Wang, 1990), что Ectopocynus принадлежит к Hesperocyoninae.
В то же время имеющийся ограниченный материал не позволяет более точно оценить филогенетическое положение Ectopocynus внутри Hesperocyoninae. Никакая синапоморфность не объединяет его с каким-либо другим конкретным родом, хотя некоторые некоторые характеристики, связанные с нижней челюстью и зубами (например, потерей метаконидов, потерей p1 и m3 и т. д.) также обнаруживаются в прогрессивных видах Enhydrocyon.
Ectopocynus simplicidens
ГОЛОТИП: RAM 25431, полная правая нижнечелюстная ветвь с зубным рядом.
ОБСУЖДЕНИЕ: мандибулярные экземпляры, составляющие ископаемый материал морфологически, в основном, контрастируют с Enhydrocyon.
Ectopocynus antiquus
ГОЛОТИП: F: AM 54090, частичная правая ветвь нижней челюсти с i3-cl, p2, p4-m3.
ОБСУЖДЕНИЕ: Уонг (Wang, 1990) больше обращает внимание на сравнение с типовым экземпляром некоторых других образцов, упирая на морфологию m1. Одновременно аргументированно доказывая сходство с геспероционинами.
Ectopocynus intermedius
ГОЛОТИП: FAM 27580, верхнечелюстные фрагменты с левым и правым P4-M2 и фрагмент левой нижней челюсти с p2-m3.
ОБСУЖДЕНИЕ: предполагаемая симпатрия согласуется с морфологическими доказательствами на основе морфологии коренных зубов.
ОБЩИЙ ВЫВОД: Hesperocyoninae рассматриваются различными маститыми исследователями на протяжении почти полутора веков. Последняя работа - X. Wang, 1990. Phylogenetic systematics of the Hesperocyoninae (Carnivora: Canidae). На мой приватный взгляд – лучшая попытка после статей гениальнейшего Тедфорда разобраться в этой группе.
Вывод в том, что монофилия этого подсемейства в филогенетическом анализе вовсе не поддерживается. К сожалению… И на самом деле статус как подсемейства, в основном, определяется отсутствием лишь синапоморфных характеристик в сравнении с двумя другими подсемействами: Borophaginae и Caninae.
Однако в группе наличествует некоторая эволюционная линия для монофилии Hesperocyon-Mesocyon-Enhydrocyon. К этой кладе могут также добавиться Sunkahetanka-Philotrox и сестринская клада с терминальным [i]Osbornodon.
Paraenhydrocyon в Hesperocyoninae включён условно, так как он может быть непосредственно эволюционировать от Hesperocyon напрямую.
Плохая сохранность материалов Caedocyon и Ectopocynus также пока не позволяет провести адекватную оценку их филогенетического положения… К тому же эти геспероционины – поздний арарикарий, а это уже ранний миоцен и их связь с более-менее очерченной группой Hesperocyoninae из раннего арарикария выглядит несколько зыбкой.
К тому же, коллеги, до сих пор имеется напросто некислая (простите за жаргонизм) разница в датировке олигоцен-миоценовых местонахождений североамериканских местонахождений - отсюда и идут споры в таксономической интерпретации многих образцов.
В палеонтологии отрицательный результат ещё ничего не значит.
Леон Берген
Неактивен