Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#76 05 February 2013 15:15:48
- Wilk
- Любознательный
- Откуда: Комсомольск-н/А
- Зарегистрирован: 30 November 2012
- Сообщений: 57
Re: Дельфины
Итак, "Парадокс Грея", или "С какой скоростью плавают дельфины?", или "Какова мощность мышечной системы дельфина?"
Статьи большие, но, на мой взгляд, очень интересные.
Самое главное - приводятся конкретные значения мощности и коэффициента сопротивления тела животного, это серьезная наука.
Отредактировано Wilk (05 February 2013 15:21:33)
Неактивен
#77 05 February 2013 15:17:15
- Wilk
- Любознательный
- Откуда: Комсомольск-н/А
- Зарегистрирован: 30 November 2012
- Сообщений: 57
Re: Дельфины
1. ПАРАДОКСА ГРЕЯ НЕТ.
Существует ли «парадокс Грея»?
Ж. Я. Грущанская
Журнал «Катера и Яхты» №85 (1980/3) (с сокращениями)
……………………………………………..
Серьезные попытки исследовать движение дельфина были предприняты впервые английским зоологом Джеймсом Греем в 1936 г. Наблюдая животных в открытом море, он пришел к выводу, что скорость плавания дельфинов выше, чем может позволить мощность, которой они обладают. Другими словами, он предположил существование какого-то неизвестного ученым механизма снижения гидродинамического сопротивления этих животных по сравнению с их твердой моделью или техническими объектами, которые двигаются при аналогичных условиях.
СКОРОСТЬ ДЕЛЬФИНА.
Данные современной науки фактических скоростях движения дельфинов можно подразделить на две группы. К первой относятся сведения, полученные при наблюдениях в открытом море с идущих судов. Именно таким способом Д. Грей зафиксировал скорость 10,3 м/с, а Эванс и Пауэлл - еще более высокое значение - 11,3 м/с, т. е. 40,7 км/ч.
……………………………………………………
Вторая группа данных получена в результате специально поставленных экспериментов с предварительно обученными животными. …В заливе Канеаху на Гавайских островах проводились опыты с тихоокеанским дельфином-афалиной …. Было установлено, что этот дельфин мог поддерживать максимальную скорость:
8,3 м/с - в течение 7,5 с;
7 м/с - в течение 10 с;
6,08 м/с - в течение 50 с;
3,09 м/с - в течение 24 ч.
Серия опытов была проведена в 1965 г. с двумя пелагическими дельфинами, плавающими быстрее, чем прибрежные афалины. По данным многочисленных «заплывов» (дельфины были приучены гнаться за деревянной буксируемой приманкой) удалось зарегистрировать максимальную скорость 11,06 м/с.
Автором проводился эксперимент, в котором дельфин-афалина гнался за приманкой - ярко-окрашенным шариком, двигающимся по натянутому над водой тросу. … Таким образом, максимальным дискретным значением скорости плавания дельфина может считаться величина 10-11 м/с, а среднее ее значение можно принять равным 5-б м/с.
СОПРОТИВЛЕНИЕ ДВИЖЕНИЮ.
Если методы определения сопротивления твердых тел разработаны достаточно полно, то способов непосредственного измерения сопротивления воды движению живых существ настоящее время нет. Эту задачу приходится решать косвенным путем - методом измерения сопротивления дельфина при скольжении его по инерции или так называемым методом дополнительного сопротивления.
Первый из них состоит в том, что при движении животного по инерции измеряют его мгновенные скорость и ускорение, а затем по формуле второго закона Ньютона вычисляют сопротивление.
Второй метод основывается на предположении, что хорошо тренированный дельфин развивает одну и ту же максимальную мощность при равных условиях плавания. Если животному создать какое-то строго измеренное дополнительное сопротивление, мы уменьшим его скорость при затрате той же мощности. Расчет коэффициента сопротивления дельфина производится по сопоставлению измеренных скоростей плавания дельфина в свободном движении и с дополнительным сопротивлением, которое нам известно.
……………………………………………………
Для определения изменения скорости плавания с дополнительным сопротивлением тот же дельфин был обучен проплывать то же расстояние с резиновым кольцом-поясом, надевавшимся непосредственно перед спинным плавником. Перед каждой серией пробегов. кольцо-пояс надевали на дельфина, а затем снимали. Было проведено 108 про6егов дельфина с кольцом, не считая тренировок.
Коэффициент гидродинамического сопротивления (Сх) дельфина оказался равным 0,0276, что весьма близко к значению коэффициента твердой модели дельфина Схм = 0,029 (оба значения отнесены к водоизмещению в степени 2/3; те же величины, отнесенные к площади смоченной поверхности, соответственно равны 0,00415 и 0,00435).
Это существенно важный вывод, ставящий под сомнение ряд гипотез, объясняющих высокую скорость дельфина в основном свойствами его кожного покрова.
«МОЩНОСТЬ» ДЕЛЬФИНА.
Сложнее всего оценить мощность плавающего в воде дельфина. Определив значение коэффициента Сх, зная фактическую скорость дельфина, его размеры и. следовательно, площадь смоченной поверхности, мы по принятым в судостроении простейшим формулам можем рассчитать мощность, необходимую для достижения данной скорости.
Получается, что для скоростей 10-11 м/с необходима мощность 2,8-3,6 л. с.
Так, какова же фактическая мощность дельфина? Действительно ли она в 10 раз меньше определенной нами или Грей ошибся и никакого несоответствия нет?
………..
Метод, основанный на предположении, что 1 кг мышц любого млекопитающего, в том числе дельфина и человека, способен за единицу времени произвести одинаковую работу, позволяет использовать данные экспериментов со спортсменами. Пользуясь ими, можно рассчитать мощность дельфина в зависимости от длительности ее приложения по весу его мышц. Получается, что дельфин может развивать мощность:
- в течение 1 с - 21,6 л.с.,
- в течение 6 с - 7,2 л.с.,
- в течение 1 мин - 3,6 л.с.,
- в течение суток - 0,72 л.с.
Дельфины часто выскакивают из воды вверх, прыгая «с места»: для этого они останавливаются и принимают положение носом вверх. Зная вес животного, высоту прыжка, а также время разгона по вертикали, также можно подсчитать мощность. По данным американского ученого Т. Ланга, дельфин весом 180 кг, разгоняясь за 0,7 с, выпрыгивал из воды на 1,84 м. Это соответствует мощности 6,3 л.с..
Докт. мед. наук А.3. Колчинской предложен метод расчета мощности животного по количеству расходуемого при дыхании кислорода. …… Получилось, что при движении в течение 15 с со средней скоростью 6 м/с средняя мощность животного была равна 2,4 - 3,5 л. с.
Существует, наконец, метод расчета мощности животного по кинематическим параметрам колебательных движений его движителя - хвостового плавника. Кинограммы плавания дельфина и точные аэродинамические характеристики крыла, аналогичного хвостовому плавнику, позволяют получить значение мощности 3,79 л. с. при движении со скоростью 6,7 м/с.
Как видим, определяемые совершенно разными способами значения мощности оказываются во всяком случае, не ниже, чем это необходимо (по расчету) для того, чтобы дельфин развил скорость 10-11 м/с.
Итак, можно констатировать, во-первых - фактическое совпадение коэффициента гидродинамического сопротивления живого дельфина и его твердой модели, а во-вторых - отсутствие несоответствия мощности животного и развиваемой им скорости. Другими словами, «парадокс Грея» не подтвердился.
К аналогичному выводу приходит и известный американский ученый Я.Т. Ву, который считает, что гидродинамические и физиологические характеристики дельфинов не представляют собой ничего необычного, и что коэффициенты сопротивления дельфина и твердого эллипсоида при почти полностью турбулентном обтекании близки между собой. Я. Т. Ву объясняет ошибку Грея тем, что он, оценивая мощность дельфина, брал явно заниженные значения: он принимал в расчет мощность человека, развиваемую в течение десятков минут, тогда как в течение более краткого периода времени тот же человек может развивать в несколько раз большую мощность (выше приводились соответствующие цифры для дельфина, причем кратковременная 1-секундная мощность в 30 раз выше, чем та, которую тот же дельфин способен поддерживать целые сутки!).
Однако вывод об отсутствии, «парадокса Грея» отнюдь не означает, что бесполезно изучать особенности гидродинамики дельфинов. И на сегодня одним из перспективных аспектов является исследование геометрических характеристик тел и плавников дельфинов, а также структуры обтекания их твердых моделей. Такое исследование представляет интерес как для биологов (с помощью инженеров они получают дополнительную информацию о морфологии и способе плавания этих животных), так и для инженеров (в контакте с биологами они получают возможность использования существующих в природе закономерностей для оптимизации тех или других параметров технических объектов).
Л и т е р а т у р а
Г р у ш а н с к а я Ж. Я. Методы определения скорости, гидродинамического сопротивления и мощности дельфинов. «Успехи биометрии», Т. LХХ11, вып. 5, Изд-во ЛГУ, 1975.
Г р у ш а н с к а я Ж. Я. Определение скоростей плавания дельфина в естественных условиях и расчет коэффициента его гидродинамического сопротивления. – «успехи биометрии и бионики», Т. 73, вып. 5, Изд-во ЛГУ, 1976
Отредактировано Wilk (10 February 2013 05:43:18)
Неактивен
#78 05 February 2013 15:23:03
- Wilk
- Любознательный
- Откуда: Комсомольск-н/А
- Зарегистрирован: 30 November 2012
- Сообщений: 57
Re: Дельфины
Завтра выложу еще, где доказывается, что "Парадокс" все-таки есть, и в какой мере.
Неактивен
#79 13 February 2013 11:49:36
- Wilk
- Любознательный
- Откуда: Комсомольск-н/А
- Зарегистрирован: 30 November 2012
- Сообщений: 57
Re: Дельфины
Первоисточник:
http://agarbaruk.professorjournal.ru/c/ … E-6886.pdf
Список литературы, на который проводятся ссылки по ходу - там же.
Отредактировано Wilk (13 February 2013 12:02:22)
Неактивен
#80 13 February 2013 11:56:09
- Wilk
- Любознательный
- Откуда: Комсомольск-н/А
- Зарегистрирован: 30 November 2012
- Сообщений: 57
Re: Дельфины
С сильными сокращениями, чтобы не убить тяжелой математикой, в максимально популярной форме:
АНАЛИТИЧЕСКИЕ ОБЗОРЫ. ГИДРОДИНАМИКА ДЕЛЬФИНОВ
Е.В. Романенко, доктор биологических наук, кандидат физико-математических наук
Институт проблем экологии и эволюции им. А.И. Северцева РАН
Активное изучение гидродинамики дельфинов началось с работ английского зоолога Джеймса Грея в середине тридцатых годов [1, 2]. Он обратил внимание на несоответствие скорости плавания дельфинов и запасов их мускульной энергии. Проанализировав известные ему из литературы данные о скорости плавания дельфинов и сопоставив эти данные с энергией их мускулов, Грей пришел к выводу, что запасов энергии в несколько раз меньше, чем необходимо для обеспечения наблюдаемой скорости плавания. Этот вывод вошел в научную литературу как «Парадокс Грея». Он же высказал и гипотезу для объяснения парадокса.
Грей предположил, что дельфины обладают способностью сохранять обтекание тела ламинарным при скорости плавания, когда ламинарное обтекание уже должно было бы смениться турбулентным. Он же указал возможный механизм задержки перехода ламинарного обтекания в турбулентное. Грей предположил, что на теле активно плывущего дельфина формируется благожелательный (отрицательный) градиент динамического давления, который и приводит к задержке перехода ламинарного обтекания в турбулентное. Функцию генератора отрицательного градиента динамического давления он приписывал хвостовой лопасти, отбрасывающей назад массы воды в процессе создания тяги.
……………………………….
С помощью специально разработанных приборов [27, 42, 47, 54–63] измеряли пульсации давления, скорости и касательные напряжения в различных точках пограничного слоя на теле дельфина. Установлено, что среднеквадратичная величина пульсаций давления, скорости и касательные напряжения зависят от режима плавания животного. В режиме плавания с ускорением и с постоянной скоростью, эти величины заметно меньше (в 2–3 раза), чем при движении по инерции. Измерение скорости обтекания тела дельфина одновременно на разных расстояниях от его носа вдоль тела показало наличие значительного отрицательного градиента динамического давления в режиме плавания с ускорением или с постоянной скоростью. При движении по инерции градиент динамического давления положительный.
…………………………………………
На протяжении многих лет разрабатывается гипотеза о демпфирующих способностях кожного покрова дельфинов, предложенная Крамером [65, 66]. В соответствии с этой гипотезой предполагается, что кожа дельфина способна гасить возмущения потока в пограничном слое и тем самым задерживать переход от ламинарного течения в турбулентное. Проведены многочисленные исследования морфологических и упругих особенностей кожи дельфинов [21, 67–71, 73–81]. Разрабатываются искусственные демпфирующие покрытия [82–84]. В этом направлении наблюдается несомненный успех.
Что же касается наличия предположенного механизма у дельфинов, то прямых доказательств этому нет. Более того, есть основания считать, что таким механизмом дельфины не обладают.
Действительно, рассмотрим два факта:
− Способностью быстро двигаться обладают как рыбы [85, 86], так и дельфины, несмотря на то, что в биологическом отношении они чрезвычайно отличны друг от друга, имеют различную структуру кожи, наружные покровы кожи также различны. Рыбы и дельфины имеют только одну общую черту – колебательный характер движения тела с нарастающей от головы к хвосту амплитудой, являющийся достаточным условием создания отрицательного градиента динамического давления вдоль тела и, следовательно, стабилизации обтекания.
− Сопротивление движению мертвых рыб и дельфинов, движущихся по инерции (т.е. не совершающих колебательных движений телом), не отличается от сопротивления движению жестких моделей соответствующей формы [52]. Если бы кожа дельфина обладала демпфирующими свойствами, она должна была бы работать при всех режимах плавания (в том числе и при движении по инерции).
……………………………………………………
Показано, что на теле дельфина в процессе его активного движения и особенно при больших ускорениях формируется значительный отрицательный градиент динамического давления, который заметно влияет на тонкую структуру пограничного слоя, повышает критическое число Рейнольдса перехода ламинарного обтекания в турбулентное. Впервые детально изучена кинематика хвостовой лопасти [42, 47].
Установлено, что по закону, близкому к гармоническому, изменяется не угол атаки, как до сих пор предполагалось в большинстве математических моделей, а угол наклона лопасти к горизонтальной оси. Угол атаки изменяется по весьма сложному закону и имеет величину около 3 градусов.
……………………………………………
Градиент давления оказывает влияние не только на стабилизацию ламинарного пограничного слоя, но и на тонкую структуру турбулентного. Известно, в частности, что отрицательный градиент динамического давления достаточной величины может привести даже к обратному переходу турбулентного обтекания в ламинарное [99–101] или, по крайней мере, к значительному уменьшению степени турбулентности и, следовательно, к уменьшению сопротивления трения.
…………………………………………………….
На основании полученных результатов сделан вывод о том, что турбулизация пограничного слоя на теле дельфина должна начинаться прежде всего в средней его части, там, где градиент давления близок к нулю или имеет малую положительную величину. При этом передняя и задняя части тела дельфина могут быть обтекаемы ламинарным или заметно ламинаризированным потоком. Этот вывод подтверждается наблюдениями американских ученых за плаванием дельфинов в фосфоресцирующей воде (рис. 4).
По их наблюдениям турбулентный характер обтекания действительно отчетливо проявляется именно в средней части тела животного, головная же и хвостовая части тела дельфина обтекаются значительно ламинаризированным потоком.
Приведенные теоретические выводы хорошо согласуются с полученными ранее экспериментальными данными по тонкой структуре пограничного слоя, распределению касательных напряжений и градиента динамического давления на теле активно плывущего дельфина (рис. 3). Кроме того, они хорошо объясняют особенности формы тела дельфина.
Дело в том, что угол схода тела дельфина больше критического, и поэтому его тело не является оптимальным в смысле минимизации сопротивления при ламинарном обтекании. Здесь имеется в виду в первую очередь опасность отрыва ламинарного пограничного слоя. Форма тела китообразных вообще приближается к оптимальной с точки зрения минимизации сопротивления при турбулентном обтекании [42, 47, 55]. Безотрывный характер при ламинарном обтекании обеспечивается наличием на теле животного отрицательного градиента динамического давления, теоретическая модель формирования которого изложена выше.
Таким образом, можно считать, что с учетом наличия отрицательного градиента динамического давления на теле дельфина (и, очевидно, других китообразных) форму их тела можно считать оптимальной как для ламинарного, так и для турбулентного обтекания. Это одно из наиболее важных адаптивных приспособлений, созданных природой.
Необходимо отметить, что механизм формирования на теле дельфина отрицательного градиента динамического давления оказался совсем не таким, каким его предсказал Грей [2].
Он считал, что градиент давления образуется за счет колебаний хвостовой лопасти дельфина. Анализ явления показал, что роль хвостовой лопасти в формировании отрицательного градиента динамического давления очень мала и проявляется лишь в непосредственной окрестности самой лопасти (см. кривые 7 и 8 на рис. 3). Главную же роль в этом явлении играют колебания собственно тела дельфина. Если бы у дельфина вообще отсутствовала хвостовая лопасть, но тело совершало бы колебательные движения, вдоль него все равно формировался бы градиент динамического давления. Именно этим обстоятельством можно объяснить известные факты дальних миграций китов, практически лишенных хвостовой лопасти в результате схваток с морскими хищниками.
………………………………………………………………………………………
В процессе решения задачи о механизме создания тяговых усилий хвостовой лопастью дельфина в рамках проекта РФФИ (93-04-21415) проведена ревизия [38–40] некоторых зарубежных исследований, имевших целью оценку тяги, тяговой мощности, коэффициента полезного действия хвостовой лопасти дельфина как движителя и коэффициента сопротивления его тела [3, 34–37].
В работе [3] выведена формула для оценки силы тяги, развиваемой хвостовой лопастью дельфина. За основу взято хорошо известное выражение для усредненной по времени тяги, вполне корректное для случая малоамплитудных колебаний крыла большого удлинения [103]:

Однако автор упомянутой работы (Parry D.A.) решил упростить формулу (10) путем включения некоторых кинематических параметров дельфина, которые он считал известными, в численные коэффициенты. Это привело к тому, что полученная им формула оказалась ошибочной и давала оценки тяги, более чем на порядок превышающие реальные значения.
Причина в том, что в 1949 году многие кинематические параметры хвостовой лопасти дельфина были недостаточно хорошо изучены. Например, угол атаки Parry принял равным 10 градусам, тогда как по последним данным он составляет всего около 3,3 градуса. Амплитуду колебаний хвостовой лопасти и ее смоченную поверхность Parry завысил в 2 раза. Есть еще ряд мелких неточностей.
В итоге оценки коэффициента сопротивления дельфина, выполненные Parry, оказались более чем на порядок выше значений коэффициента сопротивления, характерных (даже) для полностью турбулентного обтекания. На основании проведенных оценок были сделаны (ошибочные) выводы о значительном запасе мускульной энергии у дельфинов и об отсутствии у них каких-либо механизмов снижения гидродинамического сопротивления.
Webb [34] проводил аналогичные оценки по формуле, предложенной Parry, и пришел к тем же выводам, что и Parry. Кроме того, Webb [34], Yates [35] и Fish [37] провели оценки силы тяги дельфина на основе двумерной нестационарной теории крыла [4, 90, 91]. Для оценок параметра флюгирования они воспользовались формулой, предложенной в работе [104]
Однако Webb и Fish вместо угла наклона лопасти использовали угол атаки, который, по крайней мере, на порядок меньше угла наклона лопасти. И все вместе упомянутые авторы использовали в своих оценках формулу (11), которая пригодна лишь в случае малоамплитудных колебаний крыла (хвоста дельфина).
…………………………….
В итоге Webb и Fish получили оценки, завышенные почти на порядок, а Yates – приблизительно в три раза.
……………………………

…………………………
В проекте РФФИ № 93-04-21415 приведены результаты анализа данных о коэффициенте полезного действия движителя дельфина. Информацию на этот счет могут дать графики из работы [91], на которых, к сожалению, отсутствуют кривые для значения параметра флюгирования, равного 0,9. Тем не менее можно сказать, что коэффициент полезного действия составляет величину около 97%. По данным работы [105] коэффициент полезного действия близок к 90%. Однако в этой работе оценки сделаны для крыла, имеющего угол атаки 10 градусов, тогда как угол атаки хвостовой лопасти дельфина в 3 раза меньше. Это значит, что коэффициент полезного действия, безусловно, больше 90%. Вероятно, он заключен между 90 и 97%.
………………………………………………………..
В проекте № 99-04-48317 решены следующие задачи:
- проведена оценка тянущей и подсасывающей сил, развиваемых собственно телом дельфина, на основе соотношений, полученных в рамках малоамплитудной теории тонкого тела;
- разработана математическая модель формирования градиента динамического давления колеблющейся хвостовой лопастью дельфина;
- разработана математическая модель ускоренного движения дельфина из состояния покоя до достижения им максимальной скорости плавания с учетом полученных оценок тяги и коэффициента сопротивления.
В процессе активного плавания рыб и дельфинов тяга создается за счет локомоторной волны, распространяющейся по телу животного от головы к хвосту. При этом на основании выводов теории тонкого тела для образования тяги наиболее существенны условия движения хвостового плавника. Однако это справедливо лишь в том случае, если тело обтекается потенциальным потоком. Имеющиеся экспериментальные данные и наблюдения [55] свидетельствуют о том, что пограничный слой на теле дельфина в значительной степени турбулизирован. Иногда также наблюдается срыв вихрей со спинного плавника, грудных плавников и некоторых частей тела за миделем [102]. В этом случае изменяются условия обтекания тела и, как следствие, механизм формирования тяговых усилий. Тяга может формироваться не только хвостовой лопастью, но и собственно телом животного. Это обстоятельство необходимо иметь в виду при оценке эффективности движителей рыб и дельфинов.
………………………………………………………
........... Оценки с использованием экспериментально измеренных кинематических параметров показали, что доля тяги, развиваемая собственно телом дельфина, составляет около 25% при скорости плавания 4,3 м/с и около 10% при скорости 2,2 м/с.
Как уже упоминалось выше, английский зоолог Дж. Грей [2] предположил, что на теле активно плывущего дельфина формируется благожелательный (отрицательный) градиент динамического давления, который и приводит к задержке перехода ламинарного обтекания в турбулентное. Функцию генератора отрицательного градиента динамического давления он приписывал хвостовой лопасти, отбрасывающей назад массы воды в процессе создания тяги.
……………………………………………………………..
Вполне возможно, что обтекание собственно хвостовой лопасти набегающим потоком воды определяется формируемым в ее пределах градиентом динамического давления. Таким образом, предположение Грея о решающем влиянии отсасывающего действия хвостовой лопасти на характер обтекания тела дельфина не соответствует действительной картине. Это предположение справедливо, по-видимому, лишь по отношению к обтеканию самой лопасти.
В рамках проекта № 99-04-48317 проведена оценка предельной скорости плавания дельфина при известной развиваемой мощности [42, 47]. За основу принято известное соотношение [52], связывающее развиваемую диким животным мощность с временем плавания и массой:

Оценки предельной мощности, которую кратковременно может развивать дельфин массой 80 кг, сделанные по анализу прыжков животных, показывают, что она составляет около 3000 Вт [52]. Поэтому и в предлагаемом расчете принято, что в течение первой секунды движения дельфин массой 80 кг не может развивать мощность, большую 3000 Вт (4,08 л.с.). При дальнейшем движении развиваемую дельфином мощность можно считать уменьшающейся по закону t -1/3 в соответствии с выражением (28).
……………………………………………………………..
На рис. 13 приведена расчетная зависимость скорости движения дельфина от времени, полученные в результате численного интегрирования уравнений движения (30), (33), (35) и (37) по методу Рунге–Кутта [121].

Параметры дельфина приняты следующими: масса 80 кг, максимальная развиваемая мощность 3000 Вт, максимальная развиваемая тяга в первый момент движения равна весу тела. Известны экспериментальные данные [120], которые показывают, что предельная скорость плавания дельфина в условиях неволи составляет 11 ± 0,5 м/с, что неплохо согласуется с расчетными данными, приведенными на рис. 13.
Таким образом, способность дельфина повышать критическое число Рейнольдса и снижать степень турбулентности в пограничном слое (а, следовательно, и коэффициент сопротивления) благодаря значительному отрицательному градиенту давления позволяет ему экономно расходовать свою мощность. Правда, экономия не 7 – 8 -кратная, как предполагал Грей, а всего лишь 2-х кратная или несколько больше. Это обстоятельство, в частности позволяет хотя бы частично объяснить дальние миграции китообразных.
Результаты, полученные в процессе выполнения работ по грантам РФФИ, имеют фундаментальный характер и оказывают определяющее воздействие на развитие рассматриваемого направления в мире. Они могут быть использованы на практике при проектировании технических плавниковых движителей.
Отредактировано Wilk (14 February 2013 16:23:20)
Неактивен
#81 27 March 2013 23:48:53
- alazor
- Заглянувший
- Зарегистрирован: 27 March 2013
- Сообщений: 1
Re: Дельфины
Парадокс Грея решил в 2008 году американский ученый Тимоти Веи. По его версии, свойства кожи тут ни при чем, просто мышцы дельфина оказались сильнее, чем думал Грей.
“It turns out that the answer to Gray’s Paradox had nothing to do with the dolphins’ skin,” Wei said. “Dolphins can certainly produce enough force to overcome drag. The scientific community has known this for a while, but this is the first time anyone has been able to actually quantitatively measure the force and say, for certain, the paradox is solved.”
At peak performance, the dolphins produced between 300 and 400 pounds of force. Human Olympic swimmers, by comparison, peak at about 60 to 70 pounds of force.
Источник http://www.sciencedaily.com/releases/20 … 131334.htm
Неактивен
#82 20 June 2013 08:02:49
Re: Дельфины



Несколько красивых рисунков с дельфинами. Художник John Pitre. Взял из галереи http://art-assorty.ru/12-dzhon-pitres-john-pitre.html
Неактивен
#83 04 July 2013 17:41:24
- Crazy Zoologist
- Гость
Re: Дельфины
Косткатки и синий кит.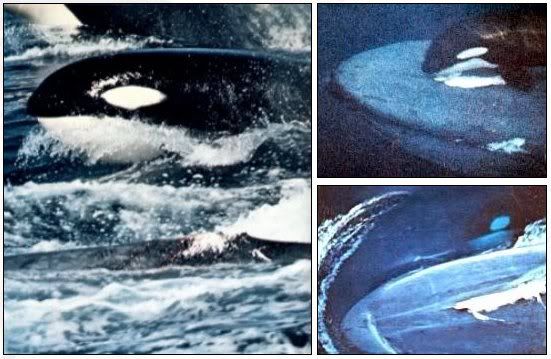
Clash of the titans: Amazing pictures of killer whales attacking pod of sperm whales twice their size on hunting expedition
By HUGO GYE
PUBLISHED: 11:58 GMT, 29 April 2013 | UPDATED: 13:27 GMT, 29 April 2013
These unique pictures show the moment a group of killer whales attacked much larger sperm whales on a relentless hunting expedition.
Five orcas pursued a pod of six sperm whales, whipping up a bloody frenzy in the middle of the ocean off the coast of Sri Lanka.
The stunning images of the hunt were captured by photographer Shawn Heinrichs, a diver who found the courage to snap away just yards from the deadly fight.
Clash: A sperm whale surfaces accompanied by two killer whales during a vicious hunt off the coast of Sri Lanka.

А это малые косатки атакуют кашалотов.
Кашалот кажется схватил косатку.

Отредактировано Кот (04 July 2013 17:45:27)
#84 18 August 2013 19:28:18
- Crazy Zoologist
- Гость
Re: Дельфины
Оказывается, дельфины иногда едят ТРАВУ!
#85 18 August 2013 21:19:37
- Вабик
- Без пяти минут зоолог

- Откуда: Великий Новгород
- Зарегистрирован: 24 May 2011
- Сообщений: 3742
Re: Дельфины
Выползают на берег за травкой?
Человек - венец природы. Терновый.
Неактивен
#86 18 August 2013 23:10:54
- Crazy Zoologist
- Гость
Re: Дельфины
Один дельфин отбирал еду у ламантина в неволе и ел ее.
#87 18 August 2013 23:36:51
- Вабик
- Без пяти минут зоолог
- Откуда: Великий Новгород
- Зарегистрирован: 24 May 2011
- Сообщений: 3742
Re: Дельфины
Может просто вредничал?))
Человек - венец природы. Терновый.
Неактивен
#88 19 August 2013 00:01:57
- Crazy Zoologist
- Гость
Re: Дельфины
Тем не менее - ел.
#89 20 August 2013 00:05:58
- Crazy Zoologist
- Гость
Re: Дельфины
Всегда удивляло, что более бочкообразные морские свиньи быстрее более обтекаемых дельфинов.
Некоторые данные о максимальной скорости передвижения некоторых видов китообразных:
Правда, немного противоречат сами себе.
И немного анатомии:

#90 05 September 2013 21:37:54
- Crazy Zoologist
- Гость
Re: Дельфины
Косатка:
#91 14 September 2013 20:14:55
- Crazy Zoologist
- Гость
Re: Дельфины
Косатка против белой акулы http://www.youtube.com/watch?v=io69Du-7Eo4
http://www.youtube.com/watch?v=SS6NjdGLVZs здесь лучше видно.
http://www.youtube.com/watch?v=XbQ5qCJEEwc косатка убила белую акулу и вырвала у нее печень.
http://www.youtube.com/watch?v=Ok6pI1iD8PU - тут фильм целиком - Nature Shock: When Killer Whales Attack.
Отредактировано Кот (14 September 2013 20:15:14)
#92 17 January 2014 15:58:54
- Вабик
- Без пяти минут зоолог
- Откуда: Великий Новгород
- Зарегистрирован: 24 May 2011
- Сообщений: 3742
Re: Дельфины

Человек - венец природы. Терновый.
Неактивен
#93 12 March 2014 15:58:12
- Crazy Zoologist
- Гость
Re: Дельфины

Выходит дельфины умнее шимпанзе?
Краткие данные об афалине:
#94 08 July 2014 06:48:31
- Krutoy_dinozavr
- Любознательный
- Зарегистрирован: 03 April 2013
- Сообщений: 65
Re: Дельфины
Дельфины, как и люди, в своей экосистеме занимают позицию высших хищников. И, скорее всего, этим сходство их биологических и даже умственных способностей отнюдь не заканчивается.
Учеными из Пенсильванского университета определен словарь дельфинов объемом уже 1012 знаков. Выяснено, что объем их активного «вокабуляра» составляет около 14 тысяч сигналов — такое же количество слов составляет и средний словарный запас человека. В обычной повседневной жизни люди и обходятся обычно 800-1000 словами. Примерно такой вот наиболее употребительный базисный лексический слой, возможно, и описан к настоящему времени учеными и для словаря дельфинов.
По своим характеристикам сигнал дельфина аналогичен чему-то наподобие иероглифа, который означает больше, чем отдельное слово.Проведенная исследователями из Пенсильванского университета магнитно-резонансная томография мозга дельфинов показала, что у дельфинов существуют ассоциативные области коры, идентичные человеческим и, по мнению специалистов: «Этот факт косвенно указывает на то, что дельфины могут быть разумными». Теменная, или двигательная, доля мозга дельфина превышает по своей площади теменную и лобную долю человека, вместе взятые. При этом затылочные зрительные доли у дельфинов чрезвычайно велики, хотя они и не особенно полагаются на зрение. Возможно, здесь имеет место кинестезия слуха и зрения - дельфины как бы «видят» с помощью слуха, испуская ультразвуки.
Они имют два органа слуха: один — обычный, другой — ультразвуковой. При этом слух дельфинов и косаток в 400-1000 раз острее человеческого.Акустическая линза на голове дельфина фокусирует ультразвук, направляя его на различные объекты. Благодаря этому дельфин и «видит» ушами. Он «ощупывает» подводный объект, определяя его форму.
Фактически, здесь можно провести аналогию с одной из наиболее важных для развития сознания человека особенностей — умением манипулировать конечностями и, особенно, мелкой моторикой рук. Дельфины также умеют «ощупывать» с помощью ультразвука. Причем получаемый таким способом образ более целостен, чем образ предмета, ощупываемого человеком руками.
Ученые провели серию экспериментов, доказывающих, что дельфины могут обмениваться весьма сложными сообщениями. Вот лишь один из примеров. Дельфину давали некое задание, которое должен был выполнить его собрат, находящийся в соседнем вольере. Через стенку вольера один дельфин «говорил» другому, что надо сделать. Например, взять красный треугольник и отдать человеку. Оба дельфина получали в награду по рыбешке. Однако было видно, что работают они не за вознаграждение, их увлекал сам процесс творческого экспериментирования. Исследователи провели тысячи опытов, задания постоянно менялись, и ни разу дельфины не ошиблись. Единственно возможный вывод из этого: дельфины прекрасно понимают все происходящее и ориентируются в мире подобно людям.
Головной мозг дельфинов и морских свиней, которые по размерам и весу соотносимы с человеком, весит несколько больше, чем у человека — до 1700 г. и имеет больше извилин. Например, еще Джон К. Лилли, много изучавший дельфинов, утверждал, что интеллект дельфина можно сопоставлять с человеческим, так как дельфины в своем общении употребляют очень сложные речевые паттерны и потому не исключена возможность установления с ними межвидовой коммуникации и общения.
И для этого существуют нетривиальные косвенные оценки.
Один из таких критериев базируется на методах статистической обработки текстов, в частности, на двух законах Джорджа Кингслева Зипфа (George K. Zipf; 1902-1950). Согласно первому закону («ранг-частота») при ранжировании частоты вхождения данного термина в документы вероятность встретить произвольно выбранное слово равно отношению количества вхождения этого слова к общему числу слов в тексте. Согласно второму закону («количество-частота») произведение частоты вхождения слова на количество разных слов с данной частотой есть величина постоянная для каждого естественного языка, которая может быть выражена функций и отображена на графике, принципиально одинаковом для любого человеческого языка. Если проранжировать слова в тексте в зависимости от того, сколько раз они употребляются, то его ранг будет примерно пропорционален его частоте в минус первой степени. Любой случайный набор знаков отражается на графике принципиально иначе. Согласно объяснению Герберта Симона, по мере развития текста возникает значимый контекст, в котором уже использовавшиеся слова имеют большую вероятность появиться еще, чем другие, случайные слова.
Данная методика была впоследствии апробирована и в отношении сигналов свиста дельфинов. Этим занимался Лоренс Дойл из Института поиска внеземного разума (SETI Institute) в Маунтин-Вью, штат Калифорния, вместе со специалистами по поведению животных из Университета штата Калифорния в Дэвисе. В результате изучения сигналов свиста дельфинов был выстроен график, аналогичный параметрам человеческих языков, несущих информацию. Данная оценка дает веские основания предполагать вероятность высокого уровня внутривидовой коммуникации среди дельфинов.
Биологи, проводившие эксперименты с дельфинами, часто с удивлением замечали и отмечали, что часто подопытные сами начинали управлять ходом эксперимента и его организаторами — людьми. Энергия творческого поиска передавалась дельфинам, и они предлагали экспериментаторам усложнить и модифицировать задание, при этом ученые неожиданно для себя замечали, что становятся экспериментальной моделью для дельфинов, которые пытались поменяться с ними ролями.
Работающие с дельфинами ученые часто приходят к вопросу: так кто же кого изучает? Люди дельфинов или дельфины людей?
Зачем дельфинам необходим столь развитой язык? Такой мощный интеллект? Столь развитой мозг? Как он мог эволюционировать?
Дельфины не занимаются, подобно людям, развитием технологий. Не используют даже камни и палки, по признакам применения которых еще в начале XX века пытались понять, обладают или нет зачатками интеллекта, например, шимпанзе.
Может дельфинам достаточно и тех колоссальных возможностей, которые они имеют от природы, может быть представляя собой в совокупности цивилизацию иного рода за свою долгую историю, десятки миллионов лет, намного опередившую людей в плане индивидуального биологического развития?
Неактивен
#95 08 July 2014 06:49:32
- Krutoy_dinozavr
- Любознательный
- Зарегистрирован: 03 April 2013
- Сообщений: 65
Re: Дельфины
Вполне возможно, что они чисто на видовом, а может даже и на родовом уровне постигли дзен)))
Неактивен
#96 08 July 2014 07:00:34
- Krutoy_dinozavr
- Любознательный
- Зарегистрирован: 03 April 2013
- Сообщений: 65
Re: Дельфины
Уже давно было известно, что дельфины обладают высокоразвитым мозгом, а наблюдения и опыты, проводившиеся в последние годы в океанариях, обнаружили, что по своей способности к дрессировке они превосходят собак и прославленных морских львов и в этом отношении могут быть поставлены выше человекообразных обезьян.
Дрессированные дельфины (рис. 400) ходят в упряжке, прыгают подобно собакам в цирке через затянутый бумагой обруч, играют в баскетбол и с большой точностью забрасывают мяч в корзину, прикреплённую на высоте в полтора метра над поверхностью воды, а одной американской дрессировщице удалось обучить дельфинов танцевать под музыку, соблюдая соответствующий ритм.
Объектом научных наблюдений и специальных физиологических экспериментов были афалины — крупные дельфины (2,3–3 м длины), водящиеся у восточных берегов Америки и у нас в Чёрном море (рис. 401).
В США изучение их поведения проводилось в особых океанариях — специально выгороженных прибрежных участках моря, достаточно просторных для свободных движений небольшой стайки дельфинов и окружённых стенами с решётками, преграждающими выход для дельфинов, но свободно пропускающими рыб и других мелких обитателей моря (в 1965 году подобный океанарий устроен и у нас на Чёрном море близ г. Судака).
Возникает вопрос: чем же мог быть обусловлен высокий уровень психической деятельности дельфинов, поставивший их в этом отношении выше всех других млекопитающих и почти рядом с человеком?
Подобно человеческим предкам (хотя и в совершенно иной обстановке), дельфины ведут стадный образ жизни, причём у них в условиях взаимного общения выработалась достаточно сложная система звуковых и сверхзвуковых сигналов. Недавние исследования показали, что вполне взрослые самцы некоторое время держатся отдельными стадами, но в основном стая дельфинов — это сильно разросшаяся семья, возглавляемая не отцом, а матерью, окружённой своими детёнышами различного возраста (дельфины-самки, как правило, рождают по одному детёнышу в год).
В таких стаях-семьях, в течение ряда лет объединяющих особей различного возраста и различного жизненного опыта, но постоянно общающихся между собой при помощи развитой сигнальной системы, и могла создаться своего рода коллективная память — возможность «устной» передачи от поколения к поколению результатов приобретаемого жизненного опыта, или, иначе говоря, возможность накопления знаний.
А так как дельфины-афалины обнаружили вдобавок и способность воспроизводить звуки человеческой речи, то некоторые исследователи (Дж. Лилли и др.) питают надежду войти в более тесный контакт с этими обитателями морей и океанов, научить их говорить по-человечески и в их лице найти не только друзей, но и толковых собеседников.
Неактивен
#97 07 August 2014 15:49:43
- Crazy Zoologist
- Гость
Re: Дельфины
Malaya kosatka pitaetsya v osnovnom molluskami (chital). No zachem togda ey takie zubi? U "obichnih" delfinov oni skromnee i ih bolshe a chelusti dlinnee, tonshe i slabee.
#98 20 August 2014 12:42:39
- Ириша
- Профессор

- Зарегистрирован: 23 February 2010
- Сообщений: 7011
Re: Дельфины
Для чего дельфины так высоко прыгают? Они наблюдают сверху таким образом или просто играют?
Как известно, бобры добры. Добротою бобры полны. Если хочешь себе добра, надо просто позвать бобра. Лишь подумай, дружок, о бобре, будешь по уши ты в добре. Если ты без бобра добр, значит, сам ты в душе бобр!
Неактивен
#99 09 February 2015 21:09:11
- Elephas Maximus
- Без пяти минут зоолог

- Откуда: Россия, Екатеринбург
- Зарегистрирован: 04 March 2012
- Сообщений: 3955
Re: Дельфины
Самый маленький бассейн для косатки в мире ![]() Тем не менее, 40 лет она протянула. https://www.youtube.com/watch?v=kmFB3_MDvCA
Тем не менее, 40 лет она протянула. https://www.youtube.com/watch?v=kmFB3_MDvCA
There are more ways than one to skin a cat.
Неактивен
#100 06 April 2015 16:34:53
- Crazy Zoologist
- Гость
Re: Дельфины
Почему косатка так поступила?![]()
https://www.youtube.com/watch?v=MFy5j4euDzQ