Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме
#126 17 April 2011 04:39:40
- Unenlagia
- Администратор

- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Файл обновлён!
Необычный гадрозавр с полым гребнем из России: азиатское происхождение ламбеозаврин.pdf
Скачать PDF
Отредактировано Unenlagia (10 June 2011 00:01:04)
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#127 27 August 2011 19:16:02
- Алекс
- Сан Саныч - Модератор

- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Кто знает, какая самая большая длина бедра азиатского зауролофа?
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#128 27 August 2011 19:32:28
- Владимир Т.
- Палеоскульптор

- Откуда: Ростов-на-Дону.
- Зарегистрирован: 14 August 2009
- Сообщений: 1765
Re: Гадрозавры
Алекс :
Кто знает, какая самая большая длина бедра азиатского зауролофа?
У голотипа (ПИН №551-8,на фото полный скелет) длинна бедра 120 см.Но есть останки более крупных особей.Вот интересно промеры ноги узнать,что на фотке. 
Моя галерея http://dinos.vx5.ru/
Неактивен
#129 27 August 2011 19:36:46
- Владимир Т.
- Палеоскульптор
- Откуда: Ростов-на-Дону.
- Зарегистрирован: 14 August 2009
- Сообщений: 1765

Неактивен
#130 27 August 2011 21:14:50
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Владимир Т. :
Примерно 150-160 см получается http://s41.radikal.ru/i092/1108/69/4a5eadd52d56t.jpg
{kind=link}
Странно что нет описания ПИНовских костяшек ,американци в этом плане не жадные .
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#131 27 August 2011 21:34:26
- Владимир Т.
- Палеоскульптор
- Откуда: Ростов-на-Дону.
- Зарегистрирован: 14 August 2009
- Сообщений: 1765
Re: Гадрозавры
Ну,эта нога по любому имеет каталожный номер и промеры-только где их взять?
Кстати,слышал на складах ПИНа монолиты лежат еще с монгольской экспедиции Ефремова.Ждут препаровки.Можно только гадать,что там интересного скрыто ![]()
Моя галерея http://dinos.vx5.ru/
Неактивен
#132 27 August 2011 22:36:12
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Владимир Т. :
Ну,эта нога по любому имеет каталожный номер и промеры-только где их взять?
Кстати,слышал на складах ПИНа монолиты лежат еще с монгольской экспедиции Ефремова.Ждут препаровки.Можно только гадать,что там интересного скрыто
Все заграничные сенсации в основном из запасников музеев , читал что есть костяхи галлилимуса размером с тарбозавра !
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#133 27 August 2011 22:49:04
- Владимир Т.
- Палеоскульптор
- Откуда: Ростов-на-Дону.
- Зарегистрирован: 14 August 2009
- Сообщений: 1765
Re: Гадрозавры
Алекс :
Владимир Т. :
Ну,эта нога по любому имеет каталожный номер и промеры-только где их взять?
Кстати,слышал на складах ПИНа монолиты лежат еще с монгольской экспедиции Ефремова.Ждут препаровки.Можно только гадать,что там интересного скрытоВсе заграничные сенсации в основном из запасников музеев , читал что есть костяхи галлилимуса размером с тарбозавра !
Это и Диновеб как то подтверждал-имеются фрагментарные останки орнитомимид размером с тарбозавра.Впринципе,ничего удивительного-есть же гигантский овираптор,да и дейнохейр явно не маленькой тварью был-руки под три метра длинной,а он к орнитомимидам еще Рождественским отнесен.
Моя галерея http://dinos.vx5.ru/
Неактивен
#134 25 September 2011 00:43:09
- Unenlagia
- Администратор
- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Новый базальный гадрозавровый динозавр из позднего мела Узбекистана и ранняя ветвь утконосых динозавров.
Ганс Дитер Сюс и Александр Аверьянов.
Levnesovia transoxiana gen. et sp. nov., из позднего мела (средний - поздний турон) Узбекистана, представляет самый старый, хорошо засвидетельствованный таксон, отнесённый к надсемейству Hadrosauroidea sensu Годфруа и др. Он отличается от несколько более раннего и близко родственного Bactrosaurus, из Внутренней Монголии (Китай), высоким сагиттальным гребнем на теменных костях и отсутствием клубо-образных поперечных отростков спинных позвонков у взрослых экземпляров. Levnesovia, Bactrosaurus и возможно Gilmoreosaurus представляют самую раннюю ветвь надсемейства Hadrosauroidea, которые обитали здесь в сеномане-туроне, а также, возможно, в Северной Америке. Вторая ветвь надсемейства Hadrosauroidea, уже сантонского возраста, включала Aralosaurus, семейство Hadrosauridae и линии, ведущие к Tanius (кампан) и Telmatosaurus (маастрихт). По всей видимости, семейство Hadrosauridae является монофилетической группой, но подсемейства Hadrosaurinae и Lambeosaurinae произошли в Северной Америке и Азии соответственно.
1. Вступление
Позднемеловые утконосые динозавры (Hadrosauridae) были одной из самых разнообразных и географически наиболее широко распространённых групп растительноядных динозавров. Их эволюционный успех был связан с серией сложных преобразований жевательного аппарата у прогрессивных орнитоподов, приведшей к более улучшенной обработке пищи (Норман и Вейсхэмпл 1985). Ранние стадии эволюции Hadrosauroidea и Hadrosauridae, по прежнему малоизвестны и предыдущие гипотезы о исторической биогеографии этих групп (Бретт-Сурмен 1979; Милнер и Норман 1984; Хэд 1998; Хэд и Кобаяши 2001) препятствовались недостатком ясных филогенетических определений и несоответствующим стратиграфическим и филогенетическим распределением критических таксонов. Здесь мы описываем самого древнего известного гадрозавроида, представляем новый филогенетический анализ прогрессивных игуанодонтовых орнитоподов, основанный на распределении 138 признаков и обсуждаем базальную ветвь надсемейства Hadrosauroidea.
2. Систематика и палеонтология
Ornithischia Seeley 1887
Ornithopoda Marsh 1881
Iguanodontia Sereno 1986
Hadrosauroidea Cope 1869 (sensu Годфруа и др. 1998)
Levnesovia transoxiana gen. et sp. nov.
Cionodon(?) kysylkumense [nomen dubium]: Рябинин 1931, p. 116.
Gilmoreosaurus arkhangelskyi [nomen dubium]: Несов 1995, p. 49.
(a) Этимология
Новый таксон назван в честь Льва Несова (1947–1995), и Transoxiana , древнее название земель за пределами реки Амударья (Oxus), соответствующих современному Узбекистану.
(b) Голотип
USNM 538191, заглазничная область крыши черепа с почти полной черепной коробкой (Рис. 1).
(с) Используемые материалы
ZIN PH 1306/16, фрагмент крыши черепа почти взрослой особи; ZIN PH 1307/16, фрагмент крыши черепа взрослой особи; USNM538126, фрагментарная черепная коробка и крыша черепа; CCMGE 565/12457, 566/12457 и ZIN PH 306/16, почти полные черепные коробки взрослых особей; USNM 538121, черепная коробка почти взрослой особи; много изолированных черепных и постчерепных костей в коллекции национального музея истории природы, Смитсоновский институт, Вашингтон, DC(USNM); центральный научно-исследовательский геологоразведочный музей имени академика Ф. Н. Чернышева, Санкт Петербург (CCMGE); зоологический институт Российской академии наук, Санкт Петербург (ZINPH); и институт зоологии, национальной академии наук Узбекистана, Ташкент (IZANUZ). Все указанные экземпляры собраны из одного местоположения и яруса (биссектинская свита, Джаракудук). Дополнительные черепные коробки и фрагменты крыши черепа, морфологически не отличимы от голотипа и оставшиеся элементы скелета, в большинстве случаев зарегистрированы многочисленными экземплярами, которые не показывают существенных таксономических отклонений и поэтому относятся к одному таксону.
(d) Местоположение и ярус
Джаракудук, Навоийская область, центральная часть пустыни Кызылкум, Узбекистан. Биссектинская свита; поздний мел (средний-поздний турон).
(e) Диагноз
Базальный гадрозавроидный игуанодонт, который отличается от известных негадрозавроидных игуанодонтов следующими признаками: отсутствует стык эктоптеригоида и скуловой кости; предзубная кость грацильная и имеет форму совка, со слегка округлённым передним краем; отверстие в надугловой кости отсутствует; коронки зуба, расположенного на зубной кости, имеют один главный гребень и слабые (если любые другие) вторичные гребни. Отличия от Bactrosaurus (Гилмор 1933) заключаются в присутствии высокого сагиттального гребня на теменной кости, отсутствии слёзного отростка на предлобной кости и отсутствии у взрослых особей клубо-образных поперечных отростков на дорсальных позвонках. Отличия от Gilmoreosaurus (Бретт-Сурмен 1979) заключаются в более коротком стыке лобной кости с орбитальной оправой, более длинном промежутке между первым зубом на зубной кости и зубом на предзубной кости (эквивалентно более чем одной пятой длины зубного ряда) и почти вертикальном венечным отростком зубной кости. Отличается от более прогрессивных Hadrosauroidea отсутствием хорошо развитого непрерывного эктоптеригоидного гребня на верхнечелюстной кости; зубной ряд на зубной кости и задний край зубной кости заканчивается на вершине венечного отростка; а также зубной ряд на зубной кости изогнут лингвально с окклюзионного вида.
(f) Комментарии
Высокий сагиттальный гребень также присутствует у негадрозаврового гадрозавроида Tanius из кампана (?) Шаньдуна, Китай (Уиман 1929) и ламбеозавринового гадрозаврида Amurosaurus из маастрихта дальневосточной России (Годфруа и др. 2004b). Этот признак отображает аутапоморфию Amurosaurus, но очевидно был развит независимо у этих трёх таксонов. Присутствие выступающего серединного отростка между базиптеригоидными отростками, Годфруа и др. (2004b) отображают как дополнительную аутапоморфию Amurosaurus. Подобный отросток присутствует у Levnesovia. Однако отсутствие этого отростка у других гадрозавроидных таксонов, скорей всего может быть случайно сохранившимся признаком, чем филогенетически важным.
Рис. 1. Levnesovia transoxiana gen. et sp. nov., частичный череп USNM 538191 (голотип), в (a) вентральном, (b) дорсальном, (c) левом латеральном виде и (d ) виде сзади. Джаракудук, центральная часть пустыни Кызылкум, Узбекистан; биссектинская свита, верхний мел (турон). Сокращения: bptp, базиптеригоидный отросток; Exo, боковая затылочная кость; fnf, лобно-носовой родничок; fo, овальное окно; Fr, лобная кость; jv, отверстие ярёмной вены; mp, серединный отросток; oc, затылочный мыщелок; Pa, теменная кость; pcr, теменной гребень; Po, заглазничная кость; pocp, завертлужный отросток; popr, зазатылочный отросток; Ps, парасфеноид; qc, вертлужная впадина квадратной кости; So, верхняя затылочная кость; Sq, чешуйчатая кость. Римскими цифрами обозначены черепные нервы. Масштабная линия, 10см.
3. Описание
(a) Череп
Предчелюстные, носовые и слёзные кости неизвестны. Плоская предлобная кость находится на одном уровне с окружающими элементами и недостающим отростком слёзной кости. На её орбитальной оправе, сзади расположены зубцы, указывающие на стык со свободной пальпебральной чашечкой (см. электронный дополнительный материал 1, Рис. a и b). Крепкий постеромедиальный гребнеобразный отросток входит в карманообразное углубление на лобной кости. Лобная кость плоская и в 1,3 - 1,5 раза больше в длину, чем в ширину (Рис. 1a,b). На её антеролатеральном краю присутствует глубокая ямка, для размещения предлобной кости. Фасетка носовой кости - узкая. Присутствует маленький родничок между лобными и носовыми костями. Лобная кость занимает только короткий участок орбитальной оправы, между предлобной и заглазничной костью. Короткая серединная ветвь заглазничной кости, прилегает к лобной и теменной кости. Лобная и заглазничная кость формируют непрерывное соединение с антеродорсальной частью латеросфеноида. Участок заглазничной кости в орбитальной оправе, подобный таковому на предлобной кости - имеет зубцы. Задняя ветвь заглазничной кости пролегает назад, к заднему краю отверстия надвисочной кости и частично сбоку налегает на чешуйчатую кость. Пролегающая антеровентрально, сужающаяся вентральная ветвь заглазничной кости, прилегает к скуловой кости. Слитные теменные кости удлинены в переднезаднем направлении. Короткий ромбовидный переднесерединный отросток теменных костей вклинен между лобными костями (Рис. 1b). Сагиттальный гребень низкий спереди и высокий сзади (Рис. 1d). Выделяющееся синусоидальное углубление вдоль дорсальной поверхности этого гребня, возможно приспособлено для некостного расширения костистого гребня. Треугольная передняя ветвь чешуйчатой кости прилегает к заглазничной кости. Углубление для размещения проксимальной головки квадратной кости, окружено предвертлужным и поствертлужным отростком. Плоский поствертлужный отроскок, пролегает вентролатерально, с отклоняющейся вперёд его дистальной точкой. Медиальная поверхность прилегает к околозатылочному отростку. На вентромедиальной стороне присутствует широкая, седлообразная суставная поверхность, для дорсального угла околозатылочного отростка. Медиальная ветвь чешуйчатой кости, это высокая вертикальная пластина, изогнутая вперёд к сагиттальному гребню, но не прилегающая к нему. На вентральном выступе присутствует вогнутость над нижней выступающей частью верхней затылочной кости. Верхнечелюстная кость треугольная с бокового вида, с острым дорсальным верхом, приблизительно посередине длины кости (электронный дополнительный материал рис. с). Единственная известная верхнечелюстная кость взрослой особи (USNM 538145) неполная спереди; в её сохранившейся части присутствует 23 зубных ряда, с тремя зубами в вертикальном зубном ряду. Антеродорсальный край кости углублен выделяющимся выступом предчелюстной кости. Отросток скуловой кости - короткий и шишкообразный; его боковая сторона закрыта фасеткой для скуловой кости. На латеральной поверхности три - пять больших отверстий, формирующих горизонтальный ряд. Глубокая медиальная впадина в виде расселины, для размещения дорсальной порции крыловидного мускула (Остром 1961), отделяет отросток скуловой кости от альвеолярной области верхнечелюстной кости. Эта впадина открывается спереди в альвеолярный канал, пролегающий вперёд и вбок к зубной батарее. Сзади этой впадины - морщинистый выступ, прилегающий к эктоптеригоиду. Альвеолярный край верхнечелюстной кости слегка вогнут. Передний отросток скуловой кости, короткий в дорсовентральном направлении и вероятно имел спереди тонкий треугольный выступ, который прилегал к предчелюстной кости, но не соприкасался снаружи с верхнечелюстной и слёзной костью, как показано в контактной фасетке на верхнечелюстной кости. Его дорсальная сторона имеет заметное углубление для размещения слёзной кости. Узкая медиальная нёбная фасетка расположена между слёзной фасеткой и глубокой ямкой, для скулового отростка верхнечелюстной кости (см. электронный дополнительный материал 1, Рис. d). Тонкий дорсальный (заглазничный) отросток пролегает приблизительно перпендикулярно к продольной оси кости. Задний отросток скуловой кости выше дорсовентрально и короче в переднезаднем направлении, чем передний отросток, прилегающий к квадратно-скуловой кости с её усечённой задней стороной. Квадратно-скуловая впадина квадратной кости широкая, с плохо определёнными границами (см. электронный дополнительный материал 1, Рис. e). Квадратоскуловая фасетка вокруг впадины, предполагает закрытие ‘околоквадратного’ отверстия. Нижнечелюстной мышелок квадратной кости - треугольный с дистального вида, с пролегающим в переднезаднем направлении латеральным мыщелком (см. электронный дополнительный материал 1, Рис. f и g). Задняя поверхность верхней затылочной кости резко наклонена вперёд, почти горизонтально, с серединным гребнем, пролегающим вперёд и заканчивающимся узлообразной структурой, прилегающей к теменным костям. Стык верхней затылочной и боковой затылочной кости, отмечен сильным горизонтальным гребнем, сопровождаемым вперёд щелеобразной поперечной бороздой. Выделяющиеся узлы на постеролатеральных углах верхней затылочной кости, прилегают к чешуйчатым костям. Структура черепной коробки подобна таковой у прогрессивных орнитоподов. Затылочный мыщелок имеет вертикальную суставную поверхность и сильно выпуклую вентральную сторону. Боковые затылочные кости не прилегают к базилярной части затылочной кости из дна эндокраниальной полости. Базальные бугорки имеют выделяющиеся базилярные части затылочной кости и базисфеноида, разделённые поперечной бороздой (Рис. 1a). Глубокая продольная ямка базисфеноида, расположена между бугорками. Сбоку от большого затылочного отверстия, на каждой стороне, расположена фасетка, возможно для проатланта, как у Iguanodon. Два из трёх отверстий для черепных нервов (ч.н.) X–XII, перфорируют боковую затылочную кость, сзади от заушной распорки. Блуждающий канал (для черепного нерва Х; ‘перилимфатический проток’ у авторов) открывается медиально в заушное (‘ярёмное’) отверстие и два более задних канала (ч.н. XI и ч.н. XII) открываются медиально в эндокраниальную полость. На медиальной стороне, округлое углубление размещает дивертикул продольной венозной системы над отверстием для ч.н. XII. Спереди от заушной распорки, длинная слуховая ямка разделена вертикальным межоконным гребнем в стременной ямке (впереди; овальное окно) и заушном отверстии (сзади). Заушное отверстие состоит из отдельных отверстий для ч.н. IX (дорсально) и внутренней ярёмной вены (вентрально). Дорсально к межоконному гребню, шов стыка заднеушной и предушной кости, пролегает дорсально вдоль зазатылочного отростка (ZIN PH 306/16). Медиальная стена предушной и заднеушной кости, напротив овального окна, расширяется в вестибулярную пирамиду, которая содержит полость внутреннего уха. На медиальной поверхности вестибулярной пирамиды, расположено отверстие для ч.н. VIII. На латеральной стороне предушной кости, вперёд к слуховой ямке, присутствует глубокая вертикальная борозда, вмещающая дорсальное отверстие для черепного нерва VII и вертикальное, для внутренней сонной артерии. Дальше вперёд расположено большое, круглое отверстие для тройничного нерва, вмещающее Гассеров ганглий для ч.н. V (Рис. 1c). Отдельные углубления отмечают направления V1 и V2–3. Орбитосфеноид и пресфеноид полностью оссифицированы. В этой области нейрокраниума, присутствуют общие отверстия для левого и правого ч.н. II, большое эллиптическое отверстие для ч.н. III и VI, и дорсально гораздо более маленькое, подобное расселине, отверстие для ч.н. IV. Дорсально, маленькое отверстие возможно представляет проход для серединной нёбной артерии. Пресфеноид - короткое, пластинообразное переднее расширение орбитосфеноида. Его передний конец формирует вентральный край большого прохода обонятельных луковиц, которые открываются спереди. Три маленьких отверстия вдоль шва, между пресфеноидом и орбитосфеноидом, возможно представляют выходы для передней глазничной артерии, передней черепной артерии и канала, для венозного анастамоза, соединяющего правую и левую глазничную пазуху. Внутренняя сонная артерия входит в гипофизарную ямку, через общие отверстия с нёбной ветвью ч.н. VII, под большими отверстиями ч.н. VI. И после выходит из эндокраниальной полости, через отверстия на вентральном дне. Отросток базиптеригоида постепенно сужается вентролатерально и имеет овальные суставные фасетки. Между этими отростками, длинные серединные отростки пролегают постеровентрально и заканчиваются слегка утолщённым концом. Базисфеноид переходит в переднем направлении в выделяющийся серединный шпорообразный парасфеноид.
(b) Нижнечелюстная кость
Предзубная кость менее дугообразная, чем таковая у Protohadros (Хэд 1998), с более прямым передним краем и отчётливым дорсальным и вентральным серединным отростком (см. электронный дополнительный материал 1, Рис. h и i), как на предзубной кости Bactrosaurus (Гилмор 1933, рис. 28). Медиальная пара питательных отверстий, большая и связана с венозными каналами; другие отверстия меньше и распределены вдоль всего переднего края. Самая большая известная предзубная кость (ZIN PH 466/16; см. электронный дополнительный материал 1, Рис. j и k) имеет 29 зубных рядов. Передний конец зубной кости умеренно изогнут. Присутствует короткая диастема, между зубом на предзубной кости и первым зубом на зубной кости. На латеральной стороне, серия сосудистых отверстий, вдоль стыка зубной и предзубной кости, позволят предположить о наличии кератинового клюва при жизни. Зубной ряд изогнут лингвально с дорсального вида и заканчивается только немного позади верхушки венечного отростка. Венечный отросток почти вертикальный, с выделяющейся венечной фасеткой на его медиальной стороне. Надугловая кость формирует заднюю часть венечного отростка. Отверстие в надугловой кости отсутствует. Ориентация угловой фасетки надугловой кости, показывает, что угловая кость была расположена латерально.
(с) Зубы
Зубы Levnesovia близко напоминают таковые у Bactrosaurus (Гилмор 1933; Годфруа и др. 1998). Верхнечелюстные зубы имеют ланцетовидные коронки, которые приблизительно в два раза больше в высоту, чем в мезиодистальную ширину и имеют выделяющийся медиальный киль (см. электронный дополнительный материал 1, Рис. l и m). Зубы на зубной кости имеют ромбовидные коронки, которые могут быть в три раза больше в высоту, чем в мезиодистальную ширину (см. электронный дополнительный материал 1, Рис. n и o). Медиальный киль относительно менее заметен и присутствует по крайней мере один вторичный продольный мезиальный гребень. Он как правило размещён ближе к дистальному краю коронки. Зубы верхнечелюстной и зубной кости, по краям имеют зубцы, поддерживаемые короткими гребнями.
(d) Постчерепной скелет
Шейные позвонки Levnesovia - опистоцельные, с коротким центром и суставным отростком позвонка, поднятым над уровнем неврального канала (см. электронный дополнительный материал 1, Рис. p). Полный крестец не известен, но совмещённые частичные крестцовые позвонки, позволяют предположить присутствие семи крестцовых позвонков. Ключичная кость - относительно большая, с маленьким ключичным бугорком, коротким вентральным отростком (‘крюк’) и лопаточной фасеткой, которая длиннее, чем гленоидная фасетка. Плечевая кость имеет низкий дельтопекторальный гребень, который не пролегает дальше середины диафиза кости. Её дистальные мыщелки сжаты медиолатерально. На подвздошной кости, вместо латерального отростка (антивертела), присутствует латеральный выступ. Лобковая кость имеет короткую подвздошную ножку; её передний (предлобковой) отросток имеет длинную шейку (см. электронный дополнительный материал 1, Рис. r). Маленький отросток, закрывающий сзади запирательную борозду - присутствует не постоянно. Дистальные мыщелки бедренной кости пролегают в переднем направлении, с их передними концами, закрывающими разгибательный туннель, у самых крупных экземпляров (см. электронный дополнительный материал 1, Рис. s). Фаланги пальцев задней конечности сжаты дорсовентрально и имеют форму копыта (см. электронный дополнительный материал 1, Рис. t).
Рис. 2. Филогенетические, временные и палеогеографические отношения среди прогрессивных игуанодонтов. Это дерево основано на дереве строгого консенсуса, представленным в электронном дополнительном материале 4. Таксоны, отмеченные красным - из Азии, синим - из Северной Америке, жёлтым - из Европы, фиолетовым - из Азии и Северной Америке. Узловые и конечные таксоны: 1, Hadrosauroidea; 2, Hadrosauridae; 3, Telmatosaurus; 4, Lambeosaurinae; 5, Hadrosaurinae.
4. Филогенетический анализ
Чтобы оценить филогенетическое положение Levnesovia, мы выполнили филогенетический анализ гадрозавроидных таксонов, основанный на распределении 138 признаков у 34 орнитоподных таксонов, с Hypsilophodon, используемым в качестве корневого вида. Признаки были собраны более чем из 20 опубликованных источников, с добавлением нескольких новых признаков (см. полный список признаков и признако - таксономическую матрицу в электронных дополнительных материалах 2 и 3). Матрица с получившимися данными была проанализирована с использованием программы NONA v. 2.0 (Голобофф 1999), запущенной под управлением WINCLADA v. 1.00.08 (Никсон 1999). Признаки со многими состояниями рассматривались, как неупорядоченные. Одна тысяча повторений экономного пошагового алгоритма (island hopper) произвели четыре самых экономных дерева, каждое длиной в 287 шагов, индексом последовательности 0,54 и индексом задержки 0,86. Дерево строгого консенсуса показано в изменённой форме на рис. 2 и полностью в электронном дополнительном материале 4.
Рис. 3. Палеогеографическое распространение прогрессивных игуанодонтов в течении (a) позднего альба - сеномана, представленное сеноманом, 95 млн. лет назад; (b) турона - коньяка, представленное туроном 90 млн. лет назад; и (с) сантоном, 85 млн. лет назад. Protohadros показан оранжевым цветом, негадрозавридные гадрозавроиды - розовым, гадрозаврины - синим и ламбеозаврины - зелёным. Смотрите список мест нахождения в электронном дополнительном материале 5. Палеогеографические карты модифицированы Смитом и др. (1994).
5. Базальная ветвь надсемейства Hadrosauroidea
В этой работе мы используем узловое определение таксона надсемейства Hadrosauroidea Годфруа и др. (1998): Bactrosaurus, Telmatosaurus, Hadrosauridae, их самых недавних общих предков и потомков. Серено (1997) предложил узловое определение для Hadrosauriformes (Iguanodon, Parasaurolophus, их самых ближайших общих предков и потомков) и стволовое определение таксона надсемейства Hadrosauroidea (все гадрозаврообразные, которые ближе к Parasaurolophus, чем к Iguanodon). Проблема с определением Серено, надсемейства Hadrosauroidea, заключается в том, что недавние исследования, включая и это, в данной работе, были не в состоянии восстановить монофилию Iguanodontidae (или эта группа ограничивалась Iguanodon и Ouranosaurus; Хэд и Кобаяши 2001; Кобаяши и Азума 2003). По определению Серено, все более прогрессивные игуанодонты, чем Iguanodon, должны быть гадрозавроидами, включая недавно открытые таксоны, подобные Probactrosaurus из среднего мела Китая (Йу и др. 2003a,b; Годфруа и др. 2005). В нашем анализе монофилия надсемейства Hadrosauroidea (sensu Годфруа и др. 1998) обеспечивается семью ярко выраженными синапоморфиями, четыре из которых - не гомоплазии (Рис. 2; см. также электронный дополнительный материал 4). Обеспечиваемая загрузка этого узла таксона - 74 процента.
Прогрессивные игуанодонты, которые парафилетически родственны надсемейству Hadrosauroidea, известны из раннего мела Азии (Altirhinus, Equijubus, Fukuisaurus, Jinzhousaurus, Lanzhousaurus, Nanyangosaurus, Penelopognathus, Probactrosaurus, Shuangmiaosaurus) и Северной Америки (Cedrorestes, Eolambia, Planicoxa). Некоторые из этих таксонов основываются только на неполном материале и нуждаются в дальнейших исследованиях. Самые прогрессивные из этих игуанодонтов, это Protohadros из сеномана Техаса (Хэд 1998). Это может свидетельствовать о том, что надсемейство Hadrosauroidea происходит в Северной Америке.
Наш анализ указывает на две различные ветви негадрозавридных гадрозавроидов (Рис. 2). Первая ветвь происходит в течении сеномана-турона (возможно берёт начало в позднем альбе), и в настоящее время плохо засвидетельствована. Верхнечелюстные зубы, с единственным хорошо выделяющимся серединным килем, из сеномана Франции и западной России (Рис 3a; электронный дополнительный материал 5) принадлежат любым самым ранним гадрозавроидам, или прогрессивным негадрозавроидным игуанодонтам, как Protohadros. Изолированным зубам зубной кости, с ромбовидными коронками, которые были найдены в сеномане Англии (‘Trachodon cantabrigiensis’) и Узбекистана (‘Gilmoreosaurus atavus’) - не хватает вторичных гребней. Возможно они являются самыми древними известными находками представителей надсемейства Hadrosauroidea (Рис. 3a).
Levnesovia из биссектинской свиты (средний-поздний турон) Узбекистана, самый древний известный таксон, относящийся к надсемейству Hadrosauroidea. Он тесно связан с Bactrosaurus, из свиты ирен дабасу, Внутренней Монголии, Китай (Гилмор 1933; Вейсхэмпл и Хорнер 1986; Годфруа и др. 1998), но особенно отличается от последнего присутствием высокого сагиттального гребня на теменных костях и отсутствием у взрослых особей клубо-образных поперечных отростков на дорсальных позвонках. Gilmoreosaurus из свиты ирен дабасу, вероятно также гадрозавроид, но плохо известный, поэтому он не был включён в наш анализ. Возраст свиты Ирен Дабасу, по прежнему плохо датирован. Первоначально его считали ранним мелом, или сеноманом (см. обзор Ван Иттербика и др. 2005). Позже, кампанский возраст этой свиты обычно указывался из-за неверной ссылки на Карри и Эберт (1993). Фактически же, эти авторы заключили, что свита Ирен Дабасу, расположенная в Ирен Нор, является ‘лучшей рассмотренной ранней сенонской эпохой’ (Карри и Эберт 1993, стр. 140), которая датируется коньякским ярусом. Несов (1995, 1997) и Аверьянов (2002) обсуждали свидетельства, для сравнения скопления позвоночных в Ирен Дабасу, верхнем Байн-Шире и биссектинской свите, предложив датировку Ирен Дабасу, туронским - коньякским ярусом. Ван Иттербик и др. (2005) предложили датировку свиты Ирен Дабасу, маастрихтским ярусом, основываясь на общих чертах харовых водорослей и ракушковых ракообразных из немегтской свиты Монголии. Однако мы считаем это заключение проблематичным, поскольку многие из рассмотренных таксонов микрофоссилий, имеют гораздо большие стратиграфические пределы, чем маастрихт. Напротив, туронский ярус биссектинской свиты, хорошо ограничен прослойкой, содержащей окаменелости морской страты. Единственное другое свидетельство присутствия представителей надсемейства Hadrosauroidea, датируемое туронским веком - скелет из свиты Матануска, Аляски и фрагментарные останки из свиты Морено Хилл, в Нью Мехико (Пэш и Мэй 1997; МакДональд и др. 2006; Рис.3b). Следующий самый древний известный гадрозавроид - Claosaurus, из свиты Канзаса - Ниобрара Чок, которая датируется от позднего коньякского до раннего сантонского века (Карпентер и др. 1995; Рис. 3b). В настоящее время, никакие из этих таксонов, не диагностируются более низким таксономическим уровнем.
Вторая ветвь негадрозавридных гадрозавроидов, произошла в течении сантона. Более прогрессивные гадрозавроиды, чем Bactrosaurus и Levnesovia, обладают хорошо развитым эктоптеригоидным гребнем на верхнечелюстной кости, зубной ряд на зубной кости - прямой с окклюзионного вида и заканчивается сзади от верхушки венечного отростка. Tanius из кампана (?) Китая, также может принадлежать этой ветви, но челюсти у этого таксона не известны (Уиман 1929; Рис. 2). Telmatosaurus, из маастрихта Румынии (Вэйсхэмпл и др. 1993; Рис. 2), является последним выжившим представителем этой сантонской ветви. Следующая группа надсемейства Hadrosauroidea, включает Hadrosaurinae, Lambeosaurinae и Aralosaurus, из сантона - кампана Казахстана (Рождественский 1968), обеспечивается семью ярко выраженными апоморфиями (электронный дополнительный материал 3) и имеет обеспечение 70 процентной загрузкой. Aralosaurus, ранее рассматривался самым базальным ламбеозаврином (Годфруа и др. 2004a), фактически же, у него отсутствует полый надчерепной гребень, но не смотря на это, он вероятно близок к предкам Lambeosaurinae.
Самые древние залегания останков, принадлежащих представителям подсемейства Hadrosaurinae, известны из сантона свиты Юто, в Миссисипи, США (Кэй и Рассел 1973) и поздне-сантонский, или ранне-кампанский Lophorhothon, из свиты Моревиль Чок, в Алабаме, США (Лэнгстон 1960; Рис. 3c). Самые древние залегания останков, представителей подсемейства Lambeosaurinae, обнаружены в сантонских - ранних кампанских стратах Казахстана: Yaxartosaurus и ‘Procheneosaurus’ convincens (основанный на скелете молодой особи; Рождественский 1968; Норман и Курзанов 1997; Рис. 3c). Этот палеобиогеографический образец самых ранних Hadrosaurinae и Lambeosaurinae, позволяет предположить, что данная группа произошла в Северной Америке и позднее появилась в Азии. В течении кампана и маастрихта, семейство Hadrosauridae достигло своего широчайшего распространения , включая Южную Америку и Антарктиду. И представители обоих подсемейств, присутствовали в Азии и Северной Америке.
Список Литературы:
Averianov, A. O. 2002 An ankylosaurid (Ornithischia: Ankylosauria) braincase from the Upper Cretaceous Bissekty Formation of Uzbekistan. Bull. Inst. R. Sci. Nat. Belg. Sci. Terre 72, 97–110.
Brett-Surman, M. K. 1979 Phylogeny and palaeobiogeography of hadrosaurian dinosaurs. Nature 277, 560–562. (doi:10.1038/277560a0)
Carpenter, K., Dilkes, D. W. & Weishampel, D. B. 1995 The dinosaurs of the Niobrara Chalk Formation (Upper Cretaceous, Kansas). J. Vert. Paleontol. 15, 275–297.
Cope, E. D. 1869[-1870] Synopsis of the extinct Batrachia, Reptilia and Aves of North America. Trans. Am. Phil. Soc. 14, 1–252. (doi:10.2307/1005355)
Currie, P. J. & Eberth, D. A. 1993 Palaeontology, sedimentology and palaeoecology of the Iren Dabasu Formation (Upper Cretaceous), Inner Mongolia, People’s Republic of China. Cretaceous Res. 14, 127–144. (doi:10. 1006/cres.1993.1011)
Gilmore, C. W. 1933 On the dinosaurian fauna of the Iren Dabasu Formation. Bull. Am. Mus. Nat. Hist. 67, 23–78.
Godefroit, P., Dong, Z., Bultynck, P., Li, H. & Feng, L. 1998 Sino-Belgian cooperation program ‘Cretaceous dinosaurs and mammals from Inner Mongolia’. 1. New Bactrosaurus (Dinosauria: Hadrosauroidea) material from Iren Dabasu (Inner Mongolia, P. R. China). Bull. Inst. R. Sci. Nat. Belg. Sci. Terre 68(Suppl.), 3–70.
Godefroit, P., Alifanov, V. R. & Bolotsky, Y. L. 2004a A re-appraisal of Aralosaurus tuberiferus (Dinosauria, Hadrosauria) from the Late Cretaceous of Kazakhstan. Bull. Inst. R. Sci. Nat. Belg. Sci. Terre 74(Suppl.), 139–154.
Godefroit, P., Bolotsky, Y. L. & Van Itterbeeck, J. 2004b The lambeosaurine dinosaur Amurosaurus riabinini, from the Maastrichtian of Far Eastern Russia. Acta Palaeontol. Pol. 49, 585–618.
Godefroit, P., Li, H. & Shang, C. 2005 A new primitive hadrosauroid dinosaur from the Early Cretaceous of Inner Mongolia (P. R. China). C. R. Palevol. 4, 697–705 (doi:10.1016/j.crpv.2005.07.004)
Goloboff, P. 1999 NONA (ver. 1.9). Software published by the author, San Miguel de Tucuma´n, Argentina. See www. cladistics.org.
Head, J. J. 1998 A new species of basal hadrosaurid (Dinosauria, Ornithischia) from the Cenomanian of Texas. J. Vert. Paleontol. 18, 718–738.
Head, J. J. & Kobayashi, Y. 2001 Biogeographic histories and chronologies of derived iguanodontians. In VII Int. Symp. on Mesozoic Terrestrial Ecosystems, pp. 107–111. Asociacio´n
Paleontologica Argentina, Publicacio´n Especial. Kaye, J. M. & Russell, D. A. 1973 The oldest record of hadrosaurian dinosaurs in North America. J. Paleontol. 47, 91–93.
Kobayashi, Y. & Azuma, Y. 2003 A new iguanadontian (Dinosauria: Ornithopoda) from the Lower Cretaceous Kitadani Formation of Fukui Prefecture, Japan. J. Vert. Paleontol. 23, 166–175.
Langston Jr, W. 1960 The vertebrate fauna of the Selma Formation of Alabama. Part 6. The Dinosaurs. Fieldiana Geol. Mem. 3, 313–363.
Marsh, O. C. 1881 Classification of Dinosauria. Am. J. Sci. ser. 3 23, 81–86.
McDonald, A. T., Wolfe, D. G. & Kirkland, J. I. 2006 On a hadrosauromorph (Dinosauria: Ornithopoda) from the Moreno Hill Formation (Cretaceous, Turonian) of New Mexico. Bull. New Mexico Mus. Nat. Hist. Sci. 35, 277–279.
Milner, A. R. & Norman, D. B. 1984 The biogeography of advanced ornithopod dinosaurs (Archosauria: Ornithischia)—a cladistic-vicariance model. In Third Symposium on Mesozoic Terrestrial Ecosystems, Short Papers (eds W.-E. Reif & D. B. Weishampel), pp. 145–150.
Tubingen, Germany: Attempto Verlag.
Nessov, L. A. 1995 [Dinosaurs of northern Eurasia: new data about assemblages, ecology and paleobiogeography]. St Petersburg, Russia: Izdatel’stvo Sankt-Peterburgskogo Universiteta. [In Russian.]
Nessov, L. A. 1997 [Cretaceous nonmarine vertebrates of northern Eurasia] (Posthumous edn. by L. B. Golovneva & A. O. Averianov). St Petersburg, Russia: Izdatel’stvo Sankt-Peterburgskogo Universiteta. [In Russian.]
Nixon, K. C. 1999 WINCLADA (Beta), v. 0.9.9. Software published by the author, Ithaca, NY. See www. cladistics.org.
Norman, D. B. & Kurzanov, S. M. 1997 On Asian ornithopods (Dinosauria: Ornithischia). 2. Arstanosaurus akkurganensis Shilin and Suslov, 1982. Proc. Geol. Assoc. 108, 191–199. (doi:10.1016/S0016-7878(97)80027-2)
Norman, D. B. & Weishampel, D. B. 1985 Ornithopod feeding mechanisms: their bearing on the evolution of herbivory. Am. Nat. 126, 151–164. (doi:10.1086/284406)
Ostrom, J. H. 1961 Cranial morphology of the hadrosaurian dinosaurs of North America. Bull. Am. Mus. Nat. Hist. 122, 1–186.
Pasch, A. D. & May, K. C. 1997 First occurrence of a hadrosaur (Dinosauria) from the Matanuska Formation (Turonian) in the Talkeetna Mountains of south-central Alaska. In Short notes on Alaska geology (eds J. G. Clough & F. Larson), pp. 99–109. Fairbanks, AK: Alaska Department of Natural Resources.
Riabinin, A. N. 1931 [On dinosaur remains from the Upper Cretaceous of the lower parts of Amu-Darya River]. Zap. Ross. Mineralog. Obshch. 40, 114–118. [In Russian.]
Rozhdestvensky, A. K. 1968 [Hadrosaurs of Kazakhstan]. In [Upper Paleozoic and Mesozoic amphibians and reptiles of the USSR], pp. 97–141. Moscow, Russia: Nauka. [In Russian.]
Seeley, H. G. 1887 On the classification of the fossil animals commonly named Dinosauria. Proc. R. Soc. Lond. 43, 165–171. (doi:10.1098/rspl.1887.0117)
Sereno, P. C. 1986 Phylogeny of the bird-hipped dinosaurs (order Ornithischia). Natl Geogr. Res. 2, 234–256.
Sereno, P. C. 1997 The origin and evolution of dinosaurs. Annu. Rev. Earth Planet. Sci. 25, 435–489. (doi:10.1146/ annurev.earth.25.1.435)
Smith, A. G., Smith, D. G. & Funnell, B. M. 1994 Atlas of Mesozoic and Cenozoic coastlines. Cambridge, UK: Cambridge University Press.
Van Itterbeeck, J., Horne, D. J., Bultynck, P. & Vandenberghe, N. 2005 Stratigraphy and palaeoenvironment of the dinosaur-bearing Upper Cretaceous Iren Dabasu Formation, Inner Mongolia, People’s Republic of China. Cretaceous Res. 26, 699–725. (doi:10.1016/ j.cretres.2005.03.004)
Weishampel, D. B. & Horner, J. R. 1986 The hadrosaurid dinosaurs from the Iren Dabasu Fauna (People’s Republic of China, Late Cretaceous). J. Vert. Paleontol. 6, 38–45.
Weishampel, D. B., Norman, D. B. & Grigorescu, D. 1993 Telmatosaurus transsylvanicus from the Late Cretaceous of Romania: the most basal hadrosaurid dinosaur. Palaeontology 36, 361–385.
Wiman, C. 1929 Die Kreide-Dinosaurier aus Shantung. Palaeontol. Sin. C 6, 1–67.
You, H., Luo, Z., Shubin, N. H., Witmer, L. M., Tang, Z. & Tang, F. 2003a The earliest-known duck-billed dinosaur from deposits of Late Early Cretaceous age in northwest China and hadrosaur evolution. Cretaceous Res. 24, 347–355. (doi:10.1016/S0195-6671(03)00048-X)
You, H., Ji, Q., Li, J. & Li, Y. 2003b A new hadrosauroid dinosaur from the Mid-Cretaceous of Liaoning, China. Acta Geol. Sin. 77, 148–154.
Электронный дополнительный материал доступен на: http://dx.doi.org/10.1098/rspb.2009.0229 или через http://rspb.royalsocietypublishing.org.
Оригинал статьи:
Hans-Dieter Sues and Alexander Averianov. (2009). A new basal hadrosauroid dinosaur from the Late Cretaceous of Uzbekistan and the early radiation of duck-billed dinosaurs. Proceedings of the Royal Society B: Biological Sciences, 276(1667): 2549-2555.
PS: Электронный дополнительный материал можно также скачать в одном архиве здесь.
Отредактировано Unenlagia (30 September 2011 01:08:02)
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#135 30 September 2011 00:00:29
- Unenlagia
- Администратор
- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Новый базальный гадрозавровый динозавр из позднего мела Узбекистана и ранняя ветвь утконосых динозавров. pdf
Ганс Дитер Сюс и Александр Аверьянов.
Скачать
Отредактировано Unenlagia (30 September 2011 00:52:43)
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#136 14 October 2011 03:29:33
- Crazy Zoologist
- Гость
Re: Гадрозавры
Черепа эдмонтозавров из Северной Америки.
В большом разрешении тут - http://upload.wikimedia.org/wikipedia/c … skulls.png
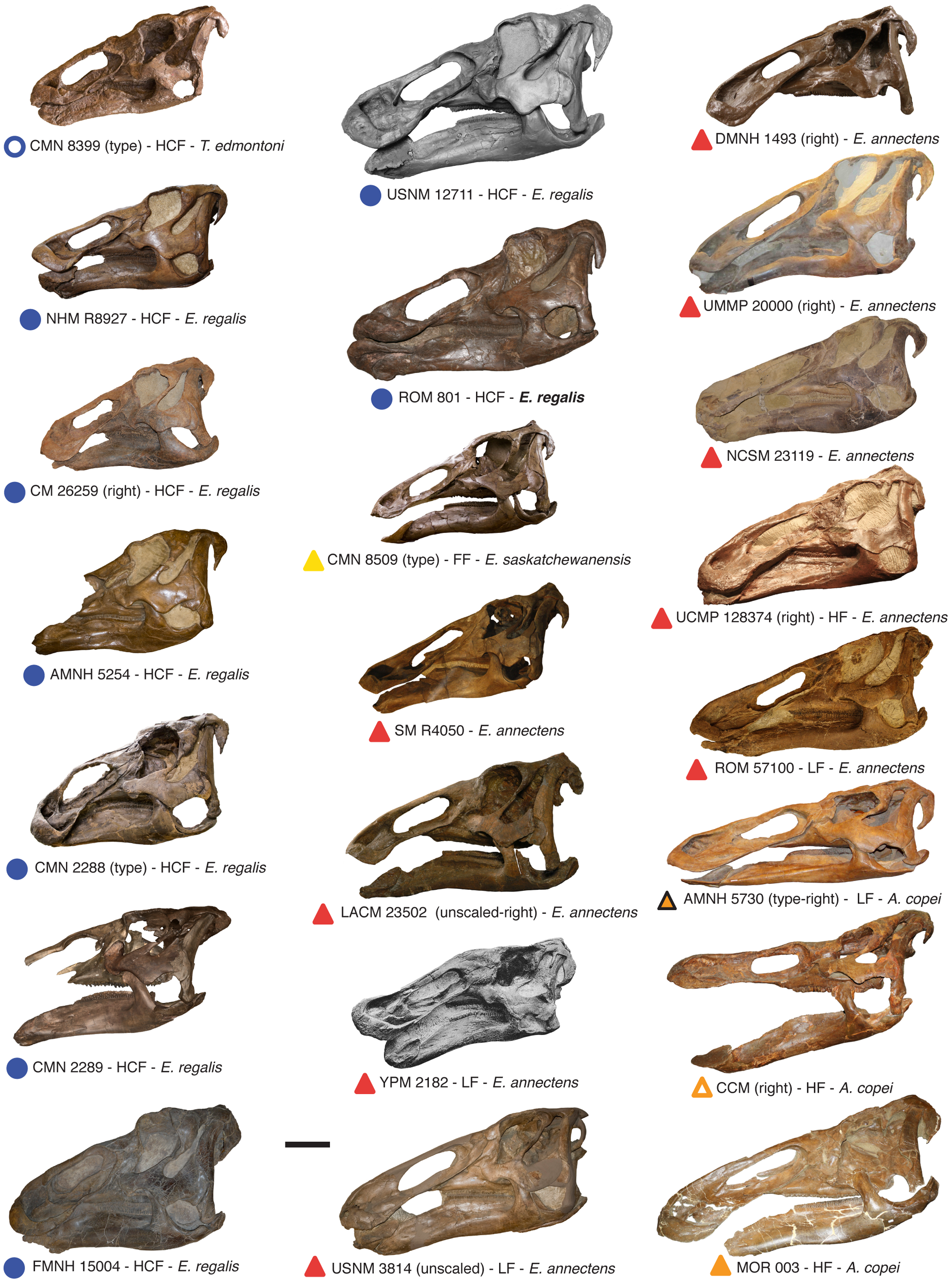{kind=link}
Отредактировано Кот (14 October 2011 03:30:45)
#137 23 November 2011 12:48:25
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
http://carnivoraforum.com/index.cgi?boa … read=13121 Huaxiaosaurus aigahtens Zhao, gen. & sp. nov. was a mild herbivorous dinosaur that lived in Zhucheng of Shandong Province in Late Mesozoic 100 Ma ago. The specimen of Huaxiaosaurus aigahtens 11.3 m in height and 18.7 m in length is a skeleton. Новый гадрозавровый динозавр ростом с брахиозавра.
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#138 23 November 2011 16:01:54
- Александр
- Любитель животных

- Откуда: Уфа
- Зарегистрирован: 28 August 2011
- Сообщений: 413
Re: Гадрозавры
Алекс :
http://carnivoraforum.com/index.cgi?board=dinosaur&action=display&thread=13121 Huaxiaosaurus aigahtens Zhao, gen. & sp. nov. was a mild herbivorous dinosaur that lived in Zhucheng of Shandong Province in Late Mesozoic 100 Ma ago. The specimen of Huaxiaosaurus aigahtens 11.3 m in height and 18.7 m in length is a skeleton. Новый гадрозавровый динозавр ростом с брахиозавра.
100млн лет назад? Получается это очень базальный представитель? И как то не верится в 11м в высоту при 18м длины - это если только ставить по-старинке, вертикально.
Гадрозавры, получается, в позднем мелу начинают, в какой то мере, занимать по всей Евразии и Северной Америке нишу зауроподов? Ведь, насколько мне известно, в северном полушарии зауроподов в мелу водилось мало, а в южном напротив, много.
PS: pdf-ка с сайта не открывается (
Отредактировано Александр (23 November 2011 16:02:55)
рисую понемногу: http://antresoll.deviantart.com/
Неактивен
#139 23 November 2011 16:03:17
- Александр
- Любитель животных
- Откуда: Уфа
- Зарегистрирован: 28 August 2011
- Сообщений: 413
Re: Гадрозавры
Чушь какая то с форумом - удалите, пожалуйста, лишние посты!
Отредактировано Александр (23 November 2011 16:03:36)
рисую понемногу: http://antresoll.deviantart.com/
Неактивен
#140 23 November 2011 18:13:02
- Unenlagia
- Администратор
- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Да, походу в этом описании он действительно признан крупнейшим орнитоподом. Длина составляет 18,7 метров, а реконструкция скелета в 11,3 метров, была сконструирована в менее реалистичной позе. Таксон представляет из себя достаточно полный скелет. Известны: мозговой отдел, левая бедренная кость, средняя часть нижней челюсти, большая часть позвоночника, плечевого пояса, передних конечностей, таза и задних ног. По поводу датировки, тут что-то пока смутно. В резюме работы действительно стоит датировка 100 млн. лет назад (т.е. по идеи - альб), однако в нидерландской википедии (статьи в остальных на данных момент вроде пока не написаны) слой формации Xingezhuangformatie, откуда известен Huaxiaosaurus, датируется кампаном. Ну и немного из описания: Череп чрезвычайно удлинённый, узкий и плоский, челюстные кости - высокие, присутствует большая зубная батарея. На позвонках шейного отдела присутствуют короткие шипообразные отростки. Передние конечности относительно короткие, с 4 пальцами и 2 м. в длину. Плечевые кости - 92/98 см. Задние конечности очень крепкие. Длина бедренной кости - 170/172 см, голень - 174/145 см.
Статья вышла на китайском: http://www.scribd.com/doc/73547424/Zhao … -1671-1688
Отредактировано Unenlagia (23 November 2011 18:28:50)
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#141 23 November 2011 18:25:11
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Unenlagia :
Да, походу в этом описании он действительно признан крупнейшим орнитоподом. Длина составляет 18,7 метров, а реконструкция скелета в 11,3 метров, была сконструирована в менее реалистичной позе. Таксон представляет из себя достаточно полный скелет. Известны: мозговой отдел, левая бедренная кость, средняя часть нижней челюсти, большая часть позвоночника, плечевого пояса, передних конечностей, таза и задних ног. По поводу датировки, тут что-то пока смутно. В резюме работы действительно стоит датировка 100 млн. лет назад (т.е. по идеи - альб), однако в нидерландской википедии (статьи в остальных на данных момент вроде пока не написаны) слой этой формации Xingezhuangformatie, датируется кампаном. Ну и немного из описания: Череп чрезвычайно удлинённый, узкий и плоский, челюстные кости - высокие, присутствует большая зубная батарея. На позвонках шейного отдела присутствуют короткие шипообразные отростки. Передние конечности относительно короткие, с 4 пальцами и 2 м. в длину. Плечевые кости - 92/98 дюймов. Задние конечности очень крепкие. Длина бедренной кости - 170/172 см, голень - 174/145 дюймов.
Статья вышла на китайском: http://www.scribd.com/doc/73547424/Zhao … -1671-1688
Саня ,не дюймов ,а сантиметров ,берцовая 147-145см. А сколько предположительно череп 155см? С высотой 11м, они явно гонят!
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#142 23 November 2011 18:34:20
- Unenlagia
- Администратор
- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Да, конечно же сантиметров, подправил. Реконструкция в 11,3 метра, как я понял, вроде бы признана некорректной, с неестественной структурой.
Отредактировано Unenlagia (23 November 2011 18:36:32)
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#143 23 November 2011 20:18:55
- Владимир Т.
- Палеоскульптор
- Откуда: Ростов-на-Дону.
- Зарегистрирован: 14 August 2009
- Сообщений: 1765
Re: Гадрозавры
У кЕтайцев,вероятно,мания-ставить гадрозавров в кенгурячью позу ![]() Вон,сколько уже манджурозавров понаделали в такой позе (к слову,у нас манджурозавр-не действительный род).
Вон,сколько уже манджурозавров понаделали в такой позе (к слову,у нас манджурозавр-не действительный род).
Завролофы из Алтан-Улы,поставленные в такую позу оценивались в 9 метров ростом-гигантизм среди гадрозаврид совсем не редкость.Вот только сильно смущает датировка альбом-череп совсем как у эдмонтозавра выглядит,то есть как у одних из последних гадрозавров.
Моя галерея http://dinos.vx5.ru/
Неактивен
#144 23 November 2011 21:27:31
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Unenlagia :
Да, конечно же сантиметров, подправил. Реконструкция в 11,3 метра, как я понял, вроде бы признана некорректной, с неестественной структурой.
Саня ,есть такие же описания скелетов Zhuchengosaurus и Shantungosaurus ?
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#145 23 November 2011 23:23:34
- Unenlagia
- Администратор
- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Самих pdf первоначальных описаний голотипов у меня нет. Но встречаются описания размеров Shantungosaurus со ссылками на эти работы. Смонтированный скелет особи средних размеров Shantungosaurus в китайском геологическом институте, Пекин, составляет 14,72 метров в длину (Glut 1997), а череп голотипа 1,63 метра (Chengzhi et al. 2001). Смонтированный скелет 16,6 метров, до выхода описания Huaxiaosaurus aigahtens, считался крупнейшим, среди птицетазовых динозавров. С дальнейшими исследованиями оба эти таксона были признаны синонимами, поскольку их апоморфные признаки различаются очень слабо. О чём более подробнее в другой статье:
Учитывая тафономические, остеологические и онтогенетические различия Zhuchengosaurus maximus Zhao и др. (2007), предположительно является синонимом Shantungosaurus giganteus Hu (1973). В этой работе также рассматривается возможное прижизненное поведение S. giganteus. Как предполагается, S. giganteus был сухопутным растительноядным динозавром, ведущим стадный образ жизни и обитавшим на холмах и долинах, в тёплом климате, с изобилием дождей и цветущей растительности.
Ji, Y., Wang, X., Liu, Y., and Ji, Q., 2011, Systematics, behavior and living environment of Shantungosaurus giganteus (Dinosauria: Hadrosauridae), Acta Geologica Sinica, 85(1): 58-65.
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#146 23 November 2011 23:34:46
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Ясно ,спасибо.
" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#147 27 December 2011 23:57:23
- Unenlagia
- Администратор
- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Четыре главных динозавровых местонахождения были исследованы в позднемеловых отложениях Амура / хэйлунцзянский регион: Цзяинь и Вулага в Китае (формация юлиангзе), Благовещенск и Кундур в России (удурчуканская свита). Более 90% найденных костей в этих локациях, принадлежат ламбеозавриновым гадрозавридам, с полыми гребнями. Charonosaurus jiayinensis из Цзяинь, Amurosaurus riabinini из Благовещенска, Olorotitan arharensis из Кундура и Sahaliyania elunchunorum из Вулаги. Плоскоголовые гадрозавриновые гадрозавриды, гораздо более малочисленны, но кажется также хорошо распространены: Kerberosaurus manakini из Благовещенска, Wulagasaurus dongi из Вулаги и новый род из Кундура. Тероподы представлены разрозненными зубами и изолированными костями; изолированные щитки и зубы, обнаруженные в Кундуре, предварительно отнесены к нодозавридам. Палинологические исследования позволяют предположить, что эти местоположения вероятно датируются одним временем с ланцианскими (северо-американский геологический ярус в верхнем мелу) скоплениями позвоночных в западе Северной Америки, которые представляют самые ранние динозавровые фауны этой окрестности. Однако, позднемеловые скопления динозавров полностью отличаются, в Амуре/хэйлунцзянский регион (ламбеозаврины изобилуют, цератопсиды отсутствуют), а в западе Северной Америки (цератопсиды изобилуют, ламбеозаврины чрезвычайно редки или отсутствуют). По всей вероятности, это отображает некий географический барьер между этими областями в маастрихтские времена, а не сильные различия в палеоэкологических условиях.
Pascal Godefroit, Pascaline Lauters, Jimmy Van Itterbeeck, Yuri L. Bolotsky & Ivan Y. Bolotsky 2011 Recent advances on study of hadrosaurid dinosaurs in Heilongjiang (Amur) River area between China and Russia. Global Geology 2011 (3)
Отредактировано Unenlagia (28 December 2011 03:24:42)
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#148 08 April 2012 00:54:49
- Unenlagia
- Администратор
- Откуда: Серпухов
- Зарегистрирован: 04 November 2008
- Сообщений: 4591
Re: Гадрозавры
Стандартизированная терминология и возможность таксономического определения гадрозаврид по отпечаткам кожи: на примере Saurolophus из Канады и Монголии.pdf
Фил Р. Белл (2012)
Скачать PDF
Оригинал статьи:
Bell P.R. (2012) Standardized terminology and potential taxonomic utility for hadrosaurid skin impressions: a case study for saurolophus from Canada and Mongolia. PLoS ONE 7(2): e31295.
Перевод Игоря Казанова, моя редакция.
Отредактировано Unenlagia (15 April 2012 03:38:55)
Не волим змајеве, али чини ми се да су ме воле!
Неактивен
#149 24 April 2012 15:44:54
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Самый крупный ламбеозаврин С/А 

" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен
#150 24 April 2012 16:21:23
- Алекс
- Сан Саныч - Модератор
- Откуда: Юг Украины
- Зарегистрирован: 05 February 2008
- Сообщений: 24254
Re: Гадрозавры
Таблицы с промерами костей гадрозаврид. 






" Из говна настоящий бульдог не получится, а вот силуэт слепить можно ". "Отрицать прошлое — это отрицать себя "
Неактивен