Вы не зашли.
Объявление
"Давайте только проявлять больше внимания, терпимости и уважения к чужому мнению — вот и всё." — Gennadius.
— О размещении изображений на форуме, О рекламе на форуме

#27 09 November 2015 07:47:59
- Звероящер
- Модератор

- Откуда: Фрязино, Московская обл.
- Зарегистрирован: 13 April 2007
- Сообщений: 14912
Re: Сфенакодонты
Наконец-то дошли руки. Шикарнейшая диссертация Фредерика Спиндлера (того самого, который ещё рисует и делает модели - http://www.frederik-spindler.de) по систематике и эволюции базальных сфенакодонтов:
Spindler F. (2015) The basal Sphenacodontia – systematic revision and evolutionary implications
Исследование представляет собой полную морфологическию и филотаксономическую ревизию базальных Sphenacodontia, обозначенных как парафилетичные "хаптодонтины" (т.е. не-сфенакодонтоидные Sphenacodontia). Ревизия включает филогенетические модели, большое количество анатомических сопоставлений и функциональные аспекты. Все таксоны заново описаны, что обеспечивает детальное изучение эволюционной стадии. Это первое исследование, которое включает в себя всех "хаптодонтин".
Основные результаты следующие:
1) Datheosaurus macrourus (гжельский ярус, Польша) и Callibrachion gaudryi (артинский ярус, Франция) являются не сфенакодонтами, а базальными, по-видимому, животноядными казеазаврами. Они являются одними из древнейших членов этой клады. Их повторное описание предоставляет первое свидетельство присутствия базальных казеазавров в Европе. Более того, эта ревизия проливает свет на форму тела базальных казеазавров, до сих пор известную лишь у молодых базальных казеид.
2) Один из древнейших известных экземпляров сфенакодонтов (московский ярус, Новая Шотландия), по-видимому, более продвинутый, чем считалось ранее. Несмотря на фрагментарность материала, он демонстрирует сходство с терапсидами, и, вероятно, относится к базальным Sphenacodontoidea.
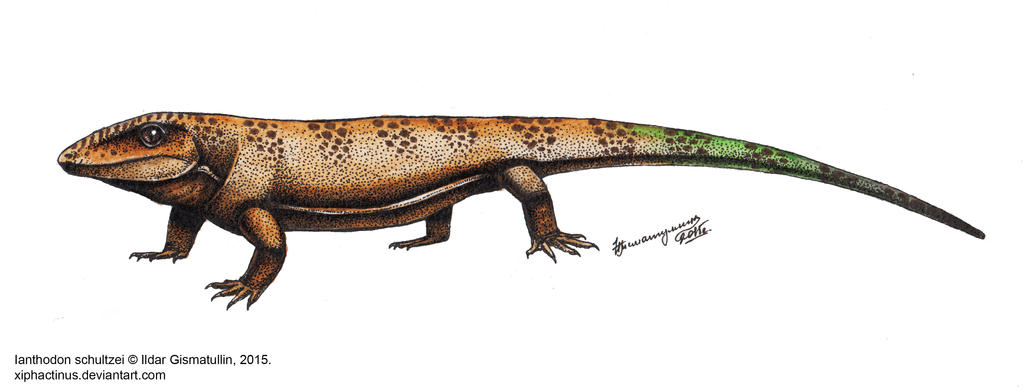
3) Ianthodon schultzei (касимовский ярус, Канзас) в настоящее время известен по значительно большему количеству материала, чем во время оригинального описания. Материал включает большую часть черепа и фрагменты посткраниального скелета. Его наиболее базальное положение среди парафилетических "хаптодонтид" хорошо обосновано. Этот вид определён как лучшая модель изначальной морфологии Sphenacomorpha (Edaphosauridae и Sphenacodontia).
4) Род Haptodus подвергся ревизии, включая ревалидацию типового вида H. baylei из артинского яруса Франции. "Haptodus" garnettensis (касимовский ярус, Канзас) не монофилетичен с Haptodus, более того, отнесённый к таксону материал демонстрирует большое разнообразие. Таким образом, он включает в себя новые таксоны Eohaptodus garnettensis gen. nov., Tenuacaptor reiszi gen. et spec. nov. и Kenomagnathus scotti gen. et spec. nov.
5) Eohaptodus garnettensis вместе с Tenuacaptor reiszi (gen. et sp. nov.), Kenomagnathus scotti (gen. et sp. nov.), базальным эдафозавридом Ianthasaurus hardestiorum и Ianthodon schultzei образуют группу с неопределенной историей признаков. Была идентифицирована адаптивная радиация с сильным указанием на трофические аннидации.
6) Крупный, но известный по фрагментарным остаткам "Haptodus" grandis (нижний цисуральский отдел, Англия) демонстрирует уникальную комбинацию признаков. Это явно более продвинутая форма, чем Haptodus и Eohaptodus, и для неё предложен новый род Hypselohaptodus gen. nov.
7) Palaeohatteria (артинский ярус, Саксония) переописана на основании повторно изученного типового материала и новых экземпляров. Кладистические ошибки, вызванные слабым окостенением у очень молодых экземпляров, могут быть исправлены с помощью тестирования реконструкций взрослых особей. Положение таксона рядом с Pantelosaurus подтверждается, и предоставлены аргументы в поддержку монофилетического статуса образуемой этими таксонами клады Palaeohatteriidae.
8) Ревизия Pantelosaurus (ассельский ярус, Саксония) выявила новые особенности в не изученных ранее частях скопления скелетов. Этот род отражает последовательное базальное продолжение тенденции, наблюдаемой у Sphenacodontidae.
9) Для Cutleria (нижний цисуральский отдел, Колорадо) не подтверждено положение среди сфенакодонтид. В отличие от всех предыдущих анализов, смесь производных и базальных признаков не позволяют сделать окончательный вывод о том, является ли Cutleria более продвинутой, чем Palaeohatteriidae.
10) Tetraceratops (верхний цисуральский отдел, Техас) признан сестринским таксоном для
Sphenacodontoidea. Фрагментарный череп этой специализированной формы не демонстрирует убедительных синапоморфий с Therapsida.
11) Для Raranimus (роадский ярус, Китай) ставится вопрос о количестве функционирующих клыков. В качестве предварительного результата таксон не признаётся стволовым представителем терапсид. Вместо этого он помещается среди базальных узлов субклад терапсид.
12) Определены три быстрые адаптивные радиации. Древнейшая из них включает Eohaptodus и его родственников. Поскольку Ianthodon может быть исключён из Sphenacomorpha, был введён более крупный стволовой таксон Haptodontiformes. Этот таксон включает Ianthodon и, вероятно, другие таксоны, относящиеся к этой радиации, но не включённые в узловой таксон Sphenacomorpha (Sphenacodontia + Edaphosauridae).
13) Более продвинутые "хаптодонтины", по-видимому, образуют вторую карбоновую радиацию. Cutleria, Tetraceratops, Hypselohaptodus, Palaeohatteriidae (Pantelosaurus и Palaeohatteria) и Sphenacodontoidea сгруппированы в таксон Pantherapsida.
14) Стратиграфический промежуток в летописи происхождения Therapsida (т.н. провал Олсона) не имеет значения. Призрачная линия в ранней перми – единственная модель, последовательно объясняющая характер эволюции и экологическую обстановку. Экологическая замена
сфенакодонтид терапсидами связана не с прямой конкуренцией, а со сдвигом биомов.
В общем, "хаптодонтины", несмотря на поверхностную схожесть, оказались очень разнообразными и неоднородными. Диссертация делает большую отсылку к гипотезе Кэмпа о происхождении терапсид (Kemp 2006):
Kemp T. S. (2006) The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis
Согласно ей, пеликозавровые сообщества наземных тетрапод были адаптированы к влажному тропическому климату. Большинство находок раннепермских пеликозавров ограничено тогдашней экваториальной зоной – областью с постоянным тёплым влажным климатом, ограниченной горами (Everwet Biome по Kemp 2006). Пеликозавров обычно сопровождает околоводная и водная фауна: различные рыбы, амфибии и базальные амниоты. Также в этих сообществах было отмечно относительно большое количество наземных хищников, и относительно мало наземных растительноядных. Возможно, позднекарбоновые и раннепермские пеликозавровые наземные сообщества ещё во многом зависели от пресноводной биоты как источника пищи, т.е. в основании пищевой пирамиды были водные растения, рыбы, различные водные тетраподы. Однако следует заметить, что пеликозавры в настоящее время не считаются настолько полуводными, как предполагалось ранее (даже такие формы, как Ophiacodon, см. Felice & Angielczyk 2014). Появление терапсид в привычном виде Кэмп связывал с аридизацией, распространением сезонного, прохладного и умеренного климата. Кэмп рассматривает три возможных сценария:
1) Гипотеза кратковременного перехода (short fuse model) – классическая. В этом случае терапсиды происходят от продвинутых сфенакодонтид в середине перми. "Преимущества" терапсид перед пеликозаврами (постановка конечностей, изменения челюстного аппарата, высокая активность, зачатки эндотермности, большая толерантность к резким колебаниям температуры, более экономное использование воды) очень быстро эволюционировали в период смены климата в середине перми. С этой гипотезой связан т.н. провал Олсона – гипотетический промежуток, в котором отсутствуют находки синапсид и где мог происходить переход от сфенакодонтов к терапсидам.
2) Гипотеза долговременного перехода (long fuse model) – терапсиды происходят от пеликозавров уровня "хаптодонтин" в результате длительного накопления ряда адаптаций и преадаптаций, позволивших к середине перми приспособиться к аридизации. Этот сценарий предпочёл Кэмп. По мнению Кэмпа, предки терапсид уже присутствовали в пеликозавровых сообществах ранней перми, но долгое время оставались незамеченными, т.к. до середины перми были почти неотличимыми от остальных пеликозавров. Комплекс экологических и биологических обстоятельств сделал возможным их обитание в граничащих с экваториальным биомом тропических регионах с сезонной влажностью (Tropical Summer Wet Biome), где в дальнейшем проходила значительная часть эволюции группы в ранней перми. Возврату в экваториальный влажный биом мешала конкуренция со стороны крупных сфенакодонтов. В середине перми изменение климата открыло для предков терапсид новые возможности заселения аридных и умеренных биомов, к которым они были приспособлены лучше пеликозавров. Также оно привело к исчезновению пеликозавровых сообществ с доминирующими крупными хищными сфенакодонтами даже без прямого взаимодействия с терапсидами.
3) Гипотеза призрачной линии (ghost lineage model) – терапсиды эволюционировали отдельно от пеликозавровых сообществ, начиная с самых ранних этапов эволюции группы. Кэмп считал этот вариант одним из наименее вероятных, аргументировав тем, что во всех известных раннепермских фаунах, обитавших вне экваториального биома, близкие к терапсидам животные не найдены.
В гипотезе Кэмпа не последнее место занимает возможная конкуренция между сфенакодонтами и предками терапсид. Однако некоторые остеологические "преимущества" терапсид, теоретически могли возникнуть и неплохо работать во влажном экваториальном биоме без каких-то климатических триггеров. Поэтому Спиндлер ставит вопрос: если пеликозавры и предки терапсид жили примерно в одних местообитаниях, и могла иметь место конкуренция между ними, то почему же смена не произошла гораздо раньше? Он предлагает немного модифицированный сценарий призрачной линии. Пеликозавры были доминирующими животными в экваториальных областях, к которым были хорошо адаптированы. Но диапазон их экологической толерантности был шире, чем считал Кэмп. Они не были строго приязаны к влажному экваториальному биому и встречались также в довольно засушливых регионах с выраженной сезонностью. На самом деле пояс влажных лесов, вероятно, исчез до установления господства пеликозавров. Терапсиды долгое время эволюционировали где-то отдельно от известной фауны пеликозавров: вдали от крупных бассейнов или в высоких широтах. Их ископаемые остатки просто не найдены, поэтому провал Олсона, в котором якобы мог происходить переход от пеликозавров к терапсидам, теряет смысл. Первые терапсиды не были конкурентами хищных пеликозавров. Конкуренция между этими двумя группами могла быть разве что на самых ранних этапах эволюции, ещё до приобретения терапсидами их отличительной морфологии. В средней перми смена климата привела к сокращению пригодных для пеликозавров местообитаний и расширению зон с аридными условиями. В какой-то момент климатического сдвига относительно широкий диапазон экологической толерантности пеликозавров оказался исчерпанным, и разнообразие группы стало снижаться. Терапсиды, наоборот, распространились вместе с сухим и сезонным климатом в те места, где их раньше не было, и впервые "внезапно" появились в палеонтологической летописи.
Таким образом, не прямая конкуренция пеликозавров с терапсидами, а сдвиг биомов в результате глобального изменения условий был главной движущей силой смены сообществ. Как минимум, две группы пеликозавров продолжали существовать в средней перми, иногда совместно с терапсидами. Есть находки, предполагающие выживание пеликозавров в конце перми и даже начале триаса (включая, возможно, и сфенакодонтов, Pineiro et al. 2003), но они фрагментарные и вполне могут принадлежать диапсидным рептилиям. Ирония в том, что в конце перми-начале триаса, вероятно, имел место похожий сценарий призрачной линии, но уже с диапсидами в качестве будущих доминантов и отодвинутыми на второй план терапсидами.
И это далеко не всё! В работе очень много интересной информации, в том числе по предполагаемой экологии пеликозавров.
Life is short, but snakes are long...
Неактивен
#28 09 November 2015 11:56:24
- Eugene
- Модератор

- Откуда: Москва
- Зарегистрирован: 12 March 2013
- Сообщений: 5053
Re: Сфенакодонты
Звероящер, спасибо!
Может быть, раз вы начали уже анализ и все прочитали, великолепно суммировали, то и об экологии пеликозавров пару анонсированных интересностей, как будет время, упомянете? ![]()
Отредактировано Eugene (09 November 2015 11:57:12)
Неактивен
#29 12 November 2015 07:58:11
- Звероящер
- Модератор
- Откуда: Фрязино, Московская обл.
- Зарегистрирован: 13 April 2007
- Сообщений: 14912
Re: Сфенакодонты
Там очень много, всего не перескажешь ![]()
Позднекарбоновое местонахождение Гарнетт в Канзасе демонстрирует уникальное и до недавнего времени скрытое разнообразие базальных синапсид. Разделение оригинального материала "Haptodus" garnettensis, по меньшей мере, на три отдельных таксона основано на пропорциях черепа и количестве зубов, в то время как общая морфология и даже форма зубов практически одинаковы. Присутствие в том же местонахождении Ianthodon и базальных эдафозаврид может подтверждать быструю диверсификацию ранних хаптодонтиформ по типу питания. Нет указаний на разделение по другим признакам и функциям, например, связанным с локомоцией. Существенные отличия касаются экологических специализаций в черепе. Подобное сосуществование близких таксонов, отличающихся деталями зубной системы и формой черепа, указывает на адаптивную радиацию, приведшую к появлению нескольких ниш животноядных синапсид с конкурентным исключением. Спиндлер сравнивает пеликозавров из Гарнетта с карбоновыми дарвиновыми вьюрками.
Экологическую нишу Ianthodon трудно определить. Возможно, он вместе с Tenuacaptor образовывал группу узкомордых хищников, питавшихся относительно мягкой добычей. Редукция числа зубов не позволяет предположить насекомоядность, т.к. последняя требует большого количества мелких зубов, либо зубцов на зубах, как это наблюдается у современных насекомоядных и большеухой лисицы (Otocyon megalotis). Kenomagnathus имел высокий массивный череп и диастему, которая концентрировала силу укуса на клыках и, возможно, крупных первых премаксиллярных зубах. Он мог быть отчасти дурофагом подобно выхухоли (Desmana moschata), в рацион которой входят ракообразные и моллюски. Большое количество зубов у Eohaptodus может быть как плезиоморфным признаком, так и специализацией. Промежуточные пропорции черепа и относительно высокое обилие в гарнеттском сообществе позволяют считать Eohaptodus генерализованной формой, возможно, ближайшей к предку сфенакоморф. Притуплённые не изогнутые зубы Ianthasaurus указывают на растительноядность. Ещё один неописанный таксон из Гарнетта, вероятно, был дурофагом и демонстрирует адаптации, лежащие за пределами известного для пеликозавров спектра.
Условия, позволившие возникнуть такой разнообразной фауне синапсид, вероятно, характеризовались низким уровнем стресса. Не известно, факторы среды или физиологические факторы оказали решающее влияние на радиацию хаптодонтиформ. Разнообразие пеннсильванских и цисуральских базальных синапсид выше, чем разнообразие диапсид. Экологически обусловленное разнообразие и разделение ниш также отражается в строении зубной системы сфенакодонтов (Brink et al. 2014, работа выше). Стратегия нетерапсидных сфенакодонтов, возможно, была примером высшего уровня K‐отбора в то время.
С Pantelosaurus и Paleohatteria связаны некоторые предположения о размножении и развитии. Учитывая гистологию костей и отсутствие каких-либо следов перестройки костной ткани, большинство экземпляров палеогаттерии, за исключением двух, являются очень молодыми животными, не прожившими даже одного года. Отсутствие в местонахождении взрослых особей предполагает, что на этом месте был "детский сад", но забота о потомстве явно отсутствовала. В противоположность этому, присутствие предполагаемого родителя известно для среднепермских варанопид. Возникает вопрос, когда эта особенность эволюционировала, и существовал ли глобальный экологический триггер, который, вероятно, повлиял также и на терапсид. Однако социальное поведение не связано с появлением новых крупных таксонов.
В известняках Нидерхслих был также обнаружен мелкий продвинутый эдафозаврид, определённый как молодой Edaphosaurus. Один из экземпляров эдафозаврид, ранее отнесённый к Palaeohatteria, является самым мелким из молодых синапсид Нидерхслиха. Поскольку там же были найдены останки маленького диадектида, было сделано предположение, что вдали берега эфемерного озера располагался "детский сад" сразу для нескольких таксонов.
Один экземпляр палеогаттерии представляет собой агрегацию скелетов двух или трёх особей. Принимая во внимание редкость палеогаттерии в формации Нидерхслих, это может быть не случайным скоплением. Хотя родительская забота отсутствовала, социальное поведение возможно для молодых животных. Эта находка может пролить свет также на скопления скелетов пантелозавра.
Оба рода близки, но по некоторым признакам в строении зубов и, возможно, когтей, пантелозавр является более специализированной формой. Практически прямые зубы Pantelosaurus, сходные с зубами Eothyrididae и Callibrachion, указывают на питание мелкой скользкой добычей, вроде беспозвоночных и личинок темноспондилов. Зубы Palaeohatteria менее специализированы. Несмотря на морфологические отличия и различное стратиграфическое распространение, всё ещё сохраняется возможность объединения этих двух родов в один. Одним из спорных и любопытных аргументом против полного таксономического объединения является прогнозирование внешнего облика взрослой особи палеогаттерии. Учитывая близость двух таксонов, пантелозавр является наиболее подходящим кандидатом для реконструкции взрослой палеогаттерии. Спиндлер составил общую модель онтогенеза представителя Paleohatteriidae, в которой ранние стадии представлены экземплярами Palaeohatteria, а более поздние – Pantelosaurus. Если наиболее крупные экземпляры пантелозавра являются половозрелыми особями, можно считать, что диаметр их тазового канала примерно соответствует диаметру откладываемых яиц. Но предположительный размер яйца, в котором могла поместиться даже уменьшенная реконструкция палеогаттерии, всё равно оказывается значительно больше диаметра тазового канала крупного пантелозавра. Спиндлер предполагает, что взрослые особи Palaeohatteria по размерам были сопоставимы со Sphenacodon.
Учитывая редкость находок "хаптодонтин", было предложено их обитание вне сохранившихся бассейнов, за исключением пеннсильванской фауны Гарнетта и межгорного бассейна Бёлен (Palaeohatteria и эдафозавриды, по крайней мере, молодые особи, жили недалеко от эфемерного озера). Hypselohaptodus, Cutleria, Tetraceratops и Palaeohatteriidae, возможно, имели промежуточный экологический диапазон, а непосредственные предки терапсид обитали на значительном расстоянии от бассейнов, на возвышенностях, в более высоких широтах. В низких широтах, в низинах вблизи сохранившихся бассейнов доминирующими наземными позвоночными были сфенакодонтиды и другие хорошо известные пеликозавры, хотя этим их распространение не ограничивалось.
Отнесение Datheosaurus и Callibrachion к казеазаврам показало, что пропорции тела начали изменяться ещё до возникновения казеид и даже до появления адаптаций к растительноядности, обозначенных зубами продвинутого типа. Модифицированный тип питания или локомоции привёл к появлению формы тела, сильно отличающейся от плезиоморфных пропорций, присущих другим субкладам базальных синапсид.
Life is short, but snakes are long...
Неактивен
#30 12 November 2015 08:26:37
- Eugene
- Модератор
- Откуда: Москва
- Зарегистрирован: 12 March 2013
- Сообщений: 5053
Re: Сфенакодонты
Звероящер, спасибо большое! Очень круто и познавательно!
Неактивен
#31 27 November 2015 09:24:55
- Звероящер
- Модератор
- Откуда: Фрязино, Московская обл.
- Зарегистрирован: 13 April 2007
- Сообщений: 14912
Re: Сфенакодонты
Голотип и единственный известный экземпляр Bathygnathus borealis, предполагаемого сфенакодонтида из нижнепермских (283-290 млн. лет, артинский ярус) красноцветных отложений Острова Принца Эдварда, Канада, – фрагмент морды с верхнечелюстным зубным рядом. Из-за фрагментарности остатков оценка систематического положения таксона с использованием кладистического анализа никогда производилась. Однако недавнее признание филогенетического использования дентальных признаков у сфенакодонтид позволяет произвести современную оценку филогенетического положения B. borealis. Результаты показывают, что B. borealis является сестринским таксоном для Dimetrodon grandis, что подтверждается следующими особенностями строения зубов: коронки с мезиальными и дистальными зубчиками, корни удлинённые, без плицидентина. Аутапоморфией B. borealis является большая лицевая часть септомаксиллы. Так как название Bathygnathus в научной литературе имеет приоритет над Dimetrodon, авторы предлагают реверсию старшинства, чтобы сохранить более знакомое и широко используемое название Dimetrodon и обеспечить универсальность, и, таким образом, выделяют новый вид Dimetrodon borealis.
Brink K.S., Maddin H.C., Evans D.C. & Reisz R.R. (2015) Re-evaluation of the historic Canadian fossil Bathygnathus borealis from the Early Permian of Prince Edward Island
Life is short, but snakes are long...
Неактивен